Дыхание и образование тепла. Процессы прорастания зерна сопровождаются интенсивным дыханием - потреблением кислорода и образованием углекислоты. Дыхание зерна при соложении представляет практический интерес с двух точек зрения: расхода питательных веществ, составляющих потерю экстрактивных веществ в производстве солода, и образования тепла при соложении.
Интенсивное дыхание зерна начинается на самых ранних стадиях замачивания.
При дыхании зерна происходит окисление углеводов до углекислоты и воды
C6H12O6 + 6О2 = 6СО2 + 6Н2О
с выделением 674 ккал тепла на 1 г-мол сахара (глюкозы), т. е. на 180 г, или 3744 кал на 1 г израсходованного в процессе дыхания сахара.
Однако при этом в зависимости от стадии процесса и степени аэрации зерна наряду с углекислотой всегда образуется некоторое количество побочных продуктов неполного окисления углеводов и продуктов сбраживания углеводов - органических кислот, спиртов и эфиров.
Благодаря этому при нормальном ращении солода всегда появляется специфический приятный запах, несколько напоминающий запах свежих огурцов.
Прорастающее зерно обладает своеобразным типом обмена веществ и при недостаточной аэрации склонно переключаться на анаэробный тип обмена веществ, образуя в качестве продуктов обмена спирт.
Но приведенной выше балансовой формуле дыхания молекулярное объемное соотношение потребленного кислорода и образованной углекислоты (т. е. дыхательный коэффициент СО2: O2) должно быть равно единице. Однако на разных стадиях замачивания зерна и соложения дыхательный коэффициент колеблется.
В начале замачивания ячменя он больше, т. е. в это время углекислоты образуется больше, чем поглощается зерном кислорода; в этом случае при использовании в качестве дыхательного материала углеводов может образоваться спирт.
В конце соложения, наоборот, дыхательный коэффициент меньше единицы, т. е. кислорода потребляется больше, чем выделяется углекислоты. В этом случае из углеводов в результате жизнедеятельности зародыша наряду с углекислотой должны образоваться органические кислоты. Действительно, во время прорастания ячменя заметно нарастает кислотность, хотя pH остается почти без изменения. Это объясняется большой буферностью экстрактов и тем, что в результате прорастания накапливаются органические кислоты с малой константой диссоциации (молочная, янтарная, яблочная и др.).
Изменение дыхательного коэффициента у прорастающего ячменя приведено в табл. 19.
Таблица 19
| Стадия проращивания
| Влажность в %
| На 100 г сухого вещества
| Дыхательный коэффициент
|
| выделилось CO2 в мл
| поглощено O2 в мл
|
| Часы от начала замачивания:
|
|
|
|
|
|
| 20,1
|
|
| 1,381
|
|
| 35,2
|
|
| 1,071
|
|
| 40,1
|
|
| 1,062
|
|
| 42,3
|
|
| 1,064
|
| Сутки ращения:
|
|
|
|
|
| первые
| 43,0
|
|
| 0,944
|
| вторые
| 42,9
|
|
| 0,979
|
| третьи
| 43,2
|
|
| 1,013
|
| четвертые
| 43,0
|
|
| 0,985
|
| пятые
| 42,8
|
|
| 0,988
|
Потери экстрактивных веществ зерна на дыхание довольно значительны, и величина их колеблется в зависимости от условий проращивания и сорта ячменя.
Размер потерь и распределение их по дням проращивания приводятся в табл. 20.
Таблица 20
| Дни проращивания
| Температура проращивания в °С
| Потери сухого вещества ячменя в %
|
| на дыхание
| на ростки
| общие
|
| Первый
| 14,5
| 1,08
|
| 1,08
|
| Второй
|
| 1,70
| 0,45
| 2,15
|
| Третий
|
| 1,99
| 0,84
| 2,83
|
| Четвертый
|
| 0,99
| 1,07
| 2,05
|
| Пятый
|
| 0,61
| 1,05
| 1.67
|
| Шестой
|
| 0,51
| 0,13
| 0,64
|
| Седьмой
|
| 0,21
| 0,02
| 0,23
|
| Всего за 7 суток
|
| 7,09
| 3,57
| 10,66
|
В связи с тем, что при солодоращении дыхательный коэффициент не равен единице, потеря сухого вещества на дыхание зерна не будет равнозначной расходу крахмала и углеводов. Поэтому расчет теплообразования во время солодоращения ячменя нельзя вести по теплотворной способности крахмала. Образование тепла будет большим или меньшим в зависимости от того, какие вещества зерна были использованы на дыхание. Оно связано также и с гидролитическими процессами, протекающими в проращиваемом зерне, гидролизом полисахаридов, белков и т. д.
Суммарное теплообразование наиболее правильно рассчитывается по теплоте сгорания исходного ячменя, полученного солода и ростков.
По экспериментальным данным, полученным в ЦНИЛБП, теплообразование отечественных ячменей во время солодоращения может быть рассчитано по потерям сухого вещества, за вычетом потерь на замачивание и ростки с пересчетным коэффициентом, равным 4283 кал тепла на 1 г потерь сухого вещества ячменя.
Биохимический состав соложеного зерна. В прорастающем зерне протекают два взаимно-противоположных, но неразрывно между собой связанных процесса: гидролиз запасных веществ эндосперма и синтез новых веществ в прорастающем зародыше, которые резко изменяют химический состав зерна.
Углеводы. Углеводы зерна изменяются, происходит накопление растворимых сахаров за счет гидролиза крахмала и гемицеллюлоз. Некоторое количество полисахаридов (гемицеллюлоз и клетчатки) вновь образуется в зародышах и солодовых ростках.
Количество крахмала в зерне уменьшается, но возрастает содержание моно- и дисахаридов. Во время солодоращения протекает сложный процесс превращения крахмала, а не только простой его гидролиз; накапливается сахароза и отчасти мальтоза. Общее содержание растворимых сахаров при этом может достигать 12%, но в среднем составляет от 7,5 до 10,5% от сухой массы солода.
Накапливающиеся в солоде сахара придают ему сладковатый вкус и обусловливают при взаимодействии с аминокислотами во время сушки солода образование ароматических и красящих веществ (меланоидинов) солода.
Накоплению сахаров во время соложения ячменя способствуют повышенная влажность эндосперма и повышенное содержание углекислоты в воздухе, при котором подавляются процессы дыхания, но усиливаются процессы гидролиза, крахмала под влиянием ферментов зерна.
Из несбраживаемых углеводов во время прорастания зерна накапливаются пентозаны и пентозы. Количество растворимых пентозанов может повышаться с 2-3% в ячмене до 12% в зеленом солоде. Образование их связано с разрушением стенок эндосперма. Сахара, входящие в состав пентозанов, легко окисляются, что способствует появлению во время сушки солода характерного аромата с оттенком печеного ржаного хлеба.
Образование аромата солода во время сушки - сложный химический процесс, связанный с окислением, разложением и конденсацией многих веществ солода. Возможно, что в процессе сушки при разложении пентозанов отщепляются пентозы, которые при дальнейшем разложении дают фурфурол и другие альдегиды и ароматические соединения, обусловливающие запах солода.
Содержание крахмала ячменя при соложении изменяется значительно. Обычно в солоде оно уменьшается по сравнению с исходным ячменем на 5-5,5% на сухую массу. Но фактически эта величина значительно больше, так как выход солода по отношению к ячменю не превышает 87-90% по сухой массе. Если принять содержание крахмала во взятом для соложения ячмене за 100%, то в полученном готовом солоде без ростков оно будет примерно 83-85%. Таким образом, расход крахмала и его превращение в растворимые углеводы солода (мальтоза, сахароза, глюкоза и т. д.) при соложении ячменя составят около 15-17%. Качестве крахмала ячменя после соложения изменяется, и он приобретает другие физические свойства. Крахмал становится без предварительной клейстеризации более доступным воздействию амилолитических ферментов. Вязкость крахмального клейстера, полученного из солодового крахмала, значительно ниже вязкости клейстера, получаемого из ячменного крахмала. Крахмальный клейстер, полученный из солодового крахмала, с йодом дает более интенсивную фиолетово-синюю окраску, в то время как крахмальный клейстер из ячменного крахмала дает с йодом чисто синюю окраску.
Накапливающиеся при соложении ячменя моносахариды и дисахариды имеют различное происхождение. Мальтоза накапливается вследствие гидролиза крахмала амилолитическими ферментами - амилазой. Сахароза образуется в результате синтезирующей деятельности зародыша, в котором она по преимуществу и находится.
Моносахариды - гексозы и пентозы образуются в солоде з результате действия ряда ферментов: мальтазы, гидролизующей мальтозу, накапливающуюся в солоде, фермента цитазы, гидролизующей гемицеллюлозы эндосперма, и др.
Цитаза - гидролитический фермент с оптимумом действия при pH 5,0-5,5 и температуре 40-45°С. Цитаза быстро инактивируется при 60°С. Под влиянием цитазы высокополимерная гемицеллюлоза сначала переводится в низкомолекулярные соединения, нерастворимые, но сильно набухающие в воде, затем в растворимые и, наконец, в конечные продукты гидролиза - моносахариды - глюкозу и пентозы. Действие цитазы изучено подробно на ксилане, в результате гидролиза которого образуется ксилоза.
Несмотря на то что представление о цитазе как о самостоятельном ферменте довольно прочно укрепилось в технологии пивоварения, все же необходимо считать, что цитолиз, т. е. растворение эндосперма, осуществляется целой группой ферментов, р. том числе протеолитическими и пектолитическими. Это объясняется тем, что в состав эндосперма, а тем более в состав межклеточного вещества, в котором заключены крахмальные клетки, входят разнообразные вещества.
Жиры и жироподобные вещества. Общее количество жира в ячмене во время соложения уменьшается, но наряду с этим увеличивается содержание жира в массе зародышей.
Резким изменениям подвергается качество жира. Жир проращенного зерна приобретает неприятный прогорклый запах вследствие омыления его и окисления жирных кислот.
С технологической точки зрения представляют особый интерес превращения фосфатидов, в состав которых входит фосфорная кислота. Количество фосфатидов во время соложения уменьшается, и содержание свободной фосфорной кислоты и ее минеральных солей увеличивается.
Кислоты и концентрация водородных ионов. Сумма кислот (летучие и нелетучие кислоты, первичные фосфаты) значительно возрастает от стадии покоящегося зерна до стадии зеленого солода. В солоде находится небольшое количество молочной, лимонной, яблочной, муравьиной, щавелевой и других кислот. Максимум образования органических кислот приходится на последние дни соложения.
Но кислотность солода в основном зависит от фосфорной кислоты, образовавшейся в результате расщепления фитина, и аминокислот, появляющихся в солоде в результате ферментативного гидролиза белков зерна.
В повышении титруемой кислотности солода главная роль принадлежит фосфатам, отщепляемым от фосфорорганических соединений под действием особой группы гидролитических ферментов - эстераз, разрывающих эфирные связи в органических соединениях.
Фитин при соложении ячменя под действием фермента фитазы распадается на инозит и фосфорную кислоту (точнее - ее кальциево-магниевую соль).
Инозит играет важную роль как ростовое вещество, ускоряющее развитие и рост дрожжей.
Фермент фитаза принадлежит к группе эстераз. Оптимум действия этого фермента лежит при температуре около 48°C и pH 5,8.
В результате всех процессов, ведущих к накоплению кислот, начальная кислотность зерна во время соложения повышается от 1,5-2,5 до 4,5-7,5 мл нормальной щелочи на 100 г зерна. Однако несмотря на значительное повышение титруемой кислотности, концентрация водородных ионов (pH) не претерпевает существенных изменений благодаря тому, что в солоде содержится большое количество кальция, магния и калия, образующих с органическими кислотами и фосфорной кислотой соли, обладающие буферными свойствами.
Белки и аминокислоты. Во время соложения ячменя азотистые вещества зерна претерпевают глубокие превращения под влиянием ферментов, расщепляющих белки эндосперма: снижается содержание нерастворимых белков и белков, растворимых в щелочи и спирте, повышается содержание водорастворимых белков и продуктов их ферментативного гидролиза - пептонов, полипептидов и аминокислот.
В среднем за время соложения около 38% азотистых веществ становятся растворимыми в воде и переходят в раствор при затирании солода, из них: в форме аминокислот - от 9 до 20% и в форме альбуминов, свертывающихся при кипячении сусла, 6-7%.
Однако приведенные величины далеко не характеризуют истинного превращения растворимых азотистых веществ при прорастании ячменя. Фактически превращению подвергается значительно большая часть азотистых веществ эндосперма. Почти в 5 раз увеличивается зародыш ячменя. Зародыш ячменя и проросток солода содержат почти в 3 раза больше азота, чем исходное зерно.
В солоде азотистые вещества проростков, включая и солодовые ростки, составляют по крайней мере 35% общего азота исходного ячменя. Эти азотистые вещества могут возникнуть только в результате синтеза, протекающего в живом зародыше засчет продуктов глубокого протеолиза белков эндосперма по крайней мере до аминокислот.
Поэтому азотистые вещества ячменя во время соложения распадаются до аминокислот не менее чем на 55%, но из них около 25-30% идет вновь на синтез и снова превращается в нерастворимые в воде белковые вещества проростка, но уже с другими свойствами и составом.
Солодовые ростки (корешки) при технологической обработке солода удаляются, по проросток ячменя, находящийся за мякинной оболочкой, остается.
Превращения белков ячменя во время соложения протекают под действием протеолитических ферментов, находящихся в ячмене и вновь образующихся в растущем зародыше.
Ферменты, расщепляющие белки, принадлежат к обширной группе гидролаз, гидролизующих сложные соединения на более простые с участием воды.
Общая схема гидролиза, осуществляемого протеолитическими ферментами, такова:
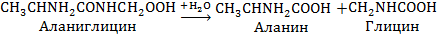
т. е. протеолитические ферменты разрывают пептидную связь. -CO-NH- и по месту разрыва присоединяют воду таким образом, что образуется карбоксильная группа одной аминокислоты и аминная группа другой. Основные ферменты, расщепляющие азотистые соединения ячменя при соложении, делятся на следующие три группы.
Протеиназы - протеолитические ферменты с оптимумом действия при pH 5,1, активируются цианидом и сульфгидрильными группами цистина и глютатиона и сероводородом. В результате действия протеиназ непосредственно на белки в процессе соложения ячменя в зерне накапливаются растворимые белки, пептоны, полипептиды и аминокислоты.
Во время соложения зерна активность протеиназ возрастает приблизительно в 40 раз.
Пептидазы - протеолитические ферменты зерна с оптимумом действия около pH 7,3-7,9. В отличие от протеиназ пептидазы не действуют непосредственно на белки зерна, а только гидролизуют пептиды, в результате чего образуются свободные аминокислоты.
Амидазы - гидролитические ферменты, катализирующие гидролиз амидов с образованием аммиака и аминокислот. Представителем этой группы ферментов являются аспарагиназа и глютаминаза. Аспарагиназа катализирует распад аспарагина на аспарагиновую кислоту и аммиак:

Глютаминаза соответственно катализирует распад глютамина.
Оптимальная зона действия их лежит в зоне pH около 8,0.
Дальнейшее превращение аминокислот - дезаминирование их в процессе обмена веществ у зародыша - осуществляется большой группой окислительных ферментов.
Пептидазы и амидазы имеют оптимум действия при таком pH, который несвойствен вытяжкам из ячменя и солода, а поэтому главную роль в гидролизе запасных веществ зерна играют протеиназы.
Таким образом, под действием этой группы ферментов во время соложения накапливаются разнообразные продукты расщепления белков: растворимые, но свертывающиеся при кипячении белки, растворимые и не свертывающиеся при кипячении белки - пептоны, полипептиды и, наконец, свободные аминокислоты.
Во время соложения аммиачный азот не накапливается ч почти не обнаруживается, аспарагин и глютамин остаются не расщепленными в солодовых ростках и эндосперме солода вследствие свойства амидаз активно катализировать распад амидов при pH 7,3-8,0 и не расщеплять их при pH 5,1-6,0.
При соложении ячменя существенное влияние на активность протеолитическнх ферментов, а следовательно, и на образование продуктов распада белков в зерне оказывают температура и степень аэрации зерна.
При повышенной температуре соложения, как правило, накапливаются главным образом водорастворимые высокомолекулярные белки. Промежуточных продуктов распада - пептонов, полипептидов, а также аминокислот - накапливается в солоде значительно меньше. Это объясняется тем, что при повышенной температуре происходит усиленный рост зародыша, благодаря чему аминокислоты, образовавшиеся при гидролизе белков эндосперма протеазами, быстро потребляются на синтез веществ растущего зародыша.
В свою очередь усиленная аэрация зерна создает неблагоприятные окислительно-восстановительные условия для действия протеиназ, и протеолиз белков идет более медленно.
Таким образом, при повышенных температурах соложения пониженное соотношение продуктов распада запасных белков эндосперма зависит от трех причин:
1) усиленного синтеза белков в зародыше,
2) быстрого потребления зародышем аминокислот и
3) торможения протеолиза вследствие неблагоприятных условий для действия протеиназ, создаваемых усиленной аэрацией.
Повышение протеолитического распада белков эндосперма во время соложения может быть достигнуто путем понижения окислительных условий, т. е. уменьшением аэрации зерна и низкой температурой ращения, путем торможения роста зародыша накапливающейся углекислотой и создания оптимальной концентрации водородных ионов (pH 5,1) внутри зерна.
О превращении белков ячменя при обычном режиме ращения солода можно судить по следующим данным (табл. 21).
Таблица 21
| Фракции азотистых соединений
| Содержание в % от общего азота
|
| в ячмене
| в солоде
|
| Аминокислоты
|
|
|
| Протеозы (средние продукты распада белка)
|
|
|
| Альбумин
|
|
|
| Глобулин
|
|
|
| Гордеин
|
|
|
| Глютелин
|
|
|
Хорошее представление о степени расщепления белков ячменя во время соложения дает определение формольного азота. Формольное число, обозначающее число миллиграммов азота, содержащегося в 100 мл 12%-ного сусла, достаточно точно характеризует степень растворения азотистых веществ. Солод, дающий формольное число свыше 230 мг, считается перерастворенным, от 200 до 230 мг - очень хорошо растворенным, от 180 до 200 мг - хорошо растворенным и до 180 мг - плохо растворенным.
О распаде белковых веществ в зеленом и сухом солоде можно судить и по содержанию растворимых и коагулируемых белков, полученных в холодной вытяжке, но лучше руководствоваться содержанием растворимых и некоагулируемых белков в конгрессном сусле. Содержание некоагулируемых белковых веществ в нем должно составлять 33%, а коагулируемые белки должны составлять не более 5%.
Прочие экстрактивные вещества. Кроме углеводов, жиров и белков, биохимическим превращениям подвергаются и другие вещества зерна, в результате чего в солоде накапливаются в свободном состоянии вещества, имеющие важное значение для технологии пивоварения и для качества пива и его питательной ценности. Отметим из них главные.
Во время соложения в солоде появляется в свободном состоянии инозит, изменяется содержание витаминов B1, B2 и аскорбиновой кислоты; содержание витамина B2 (рибофлавина) увеличивается до 210 мг % на 100 г сухого вещества солода.
Инозит, витамины B2 и B1 имеют очень важное значение для нормальной жизнедеятельности дрожжей и их бродильной активности.
Активность ферментов при соложении ячменя. Все биохимические превращения зерна во время соложения происходят под действием ферментов. Ферменты прорастающего зерна, как и всякого живого растительного организма, чрезвычайно разнообразны. Некоторые из них осуществляют все биохимические превращения, связанные с дыханием зерна и ростом зародыша. Эти ферменты важны при соложении ячменя, но для последующих технологических процессов существенного значения не имеют. Амилолитические ферменты, наоборот, имеют особое значение для технологии переработки солода и получения из него пивного сусла.
Накопление амилолитических ферментов в солоде и сохранение их в активном состоянии - одна из главных задач процесса соложения ячменя и сушки солода.
Образуются ли эти ферменты вновь или только активируются во время соложения ячменя, в данное время не важно с точки зрения технологии пивоваренного производства, хотя это и имеет большое значение для дальнейшего совершенствования процессов приготовления солода.
В ячмене и ячменном солоде содержится амилаза двух групп - α-амилаза, или декстриногенамилаза, и β-амилаза, или сахарогенамилаза.
β-Амилаза содержится в непроросшем зерне ячменя, и во время соложения ее активность увеличивается.
α-Амилаза почти полностью отсутствует в исходном ячмене и образуется в зерне в процессе соложения.
α- и β-амилазы различны но свойствам и характеру воздействия на крахмал. Эти свойства будут подробно рассмотрены ниже при изложении осахаривания солодового затора.
Амилаза в непроросшем ячмене находится в двух состояниях: в виде свободной амилазы и связанной с белком. Свободная амилаза ячменя легко переходит в раствор и активно действует на крахмал. Связанная с белком амилаза неактивна. Активность ее обнаруживается только после воздействия на зерно протеолитических ферментов.
Содержание свободной и связанной амилазы в ячмене различно. В одних сортах ячменя большая часть амилазы находится в связанном состоянии; такие ячмени при соложении требуют более глубокого протеолиза. В других, наоборот, содержится много свободной амилазы и они более легко солодятся.
Активность амилаз при проращивании ячменя показана в табл. 22 (в мг мальтозы на 100 г зерна).
Таблица 22
| Амилазная активность
| Вытяжка
|
| Водная (свободная амилаза)
| водная + папаин (общая амилаза)
|
| Непроросшего зерна
| 142,4
| 456,3
|
| Зерна после замачивания
| 79,5
| 386,7
|
| Зерна после проращивания:
|
|
|
| первый день
| 86,2
| 387,0
|
| второй день
| 111,1
| 401,9
|
| третий день
| 149,6
| 424,9
|
| четвертый день
| 211,1
| 430,9
|
| пятый день
| 285,1
| 424,9
|
| шестой день
| 276,5
| 408,6
|
Как видно из данных табл. 22, прибавление протеолитического фермента папаина при получении вытяжки из ячменя увеличивает амилазную активность более чем в 3 раза - с 142,4 до 456,3 единицы, в то время как применение папаина при получении вытяжки из солода увеличивает амилазную активность только в 1,5 раза - с 276,5 до 408,6 единицы. Это свидетельствует о том, что во время проращивания ячменя находящиеся в нем протеолитические ферменты гидролизуют белки, связывающие амилазу, и освобождают ее. Таким образом, на этом примере видно, какую важную роль играют протеолитические ферменты в активировании амилолитических ферментов в процессе соложения ячменя и насколько присутствие их может облегчить процесс осахаривания солодового затора.
В процессе соложения ячменя активность α- и β-амилазы нарастает неравномерно и не с одинаковой скоростью (рис. 32).

Во время замачивания амилолитическая активность ячменя несколько уменьшается, что, возможно, связано с выщелачиванием из ячменя солей и различных активаторов. Сильное повышение амилолитической активности наблюдается от второго до четвертого дня ращения, после чего активность повышается медленно.
Активность амилолитических ферментов в солоде и скорость ее нарастания во время соложения зависят от целого ряда условий: сорта ячменя, способа проращивания, степени аэрации и температуры процесса.
Интенсивная аэрация в начале ращения создает благоприятные условия для роста зародыша, а следовательно, и для большего накопления ферментов. Более слабая аэрация в конце соложения при уже хорошо развитом проростке, особенно при высокой концентрации углекислоты в атмосфере, усиливает протеолитический распад эндосперма, способствуя освобождению связанной амилазы.
Температура соложения играет решающую роль в накоплении ферментативной активности в солоде.
Как правило, при низких температурах соложения получается солод с наибольшей ферментативной активностью, хотя процесс соложения при этом чрезвычайно затягивается. Однако это неодинаково применимо ко всем сортам ячменя. На примере отечественных ячменей легко убедиться, что ячмени, выращенные в более жарком климате, дают удовлетворительный солод только при повышенной температуре ращения и, наоборот, ячмени, выращенные в холодном климате, как правило, резко реагируют на повышенную температуру усиленным ростом зародыша, но дают солод с недостаточной ферментативной активностью.
У солодов, полученных в результате соложения при повышенной температуре, может наблюдаться и высокая ферментативная активность, но тем не менее такие солода медленно осахариваются при изготовлении сусла. Причина такого явления кроется в том, что для быстрого осахаривания затора от солода требуется не только одна активность амилаз, но и соответственная подготовка самого крахмала солода - разрушение клеточных стенок эндосперма ферментом цитазой и белков, связывающих крахмальные зерна в эндосперме. Солод в пивоваренном производстве - это не только источник ферментов, но и сырье, на которое действуют эти ферменты. Поэтому выбор температурного режима соложения ячменя должен определяться не по какому-либо одному, а по целому ряду показателей; кроме того, он должен постоянно сочетаться с другими технологическими приемами - замачиванием ячменя и степенью аэрации зерна во время соложения.
Амилолитические ферменты в ячменном солоде распределены неравномерно. Ферментативная активность солода (в %) распределяется так:
| в верхней части эндосперма
| около 25
|
| в нижней части эндосперма
|
|
| в щитке
|
|
| в зародышевом листке
| 0,4
|
| в корешках
| 0,6
|
Амилолитическая активность зародыша (щиток, зародышевый листок и корешки) сама по себе высока, но так как масса зародыша по отношению к массе эндосперма зерна мала, то относительное количество ферментов в зародыше по отношению к общему содержанию их в зерне кажется очень небольшим.
Амилолитическая активность зародышей и эндосперма зерна (в условных единицах на 1 г сухой массы) приведена ниже.
| Часть зерна
| Общая осахаривающая активность
| Декстринирующая активность
|
| Эндосперм
| 30,08
|
|
| Зародыши
| 15,82
|
|
Декстринирующая способность зародышей почти в 4 раза больше, чем у эндосперма. Эти данные важны и интересны для технологии соложения ячменя, так как указывают на безусловное мощное образование зародышем ячменя декстринирующей амилазы (α-амилазы) во время соложения ячменя.
Величина амилолитической активности солода зависит от размера зерна. Мелкие ячмени обладают значительно большей амилолитической активностью. Такие ячмени, как правило, дают солода с большей относительной массой зародышей. Все это показывает, что зародыши ячменя играют очень важную роль в активировании ферментов, находящихся в зерне, и в образовании новых ферментов в процессе солодоращения.








