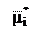Т. к. вода поглощается в осн. корневой системой, возникает необходимость ее передвижения по растению. Этот процесс составляет т. н. восходящий ток. Следует отметить, что это название отражает характер передвижения и его локализацию в растении. Он проходит по мертвым тканям стебля или черешка — сосудам у покрытосеменных и трахеидам у голосеменных. Однако эта локализация не абсолютна: вода способна перемещаться и по флоэмной системе. Передвижение по сосудам идет с большей скоростью (20 см/ч), чем по трахеидам (только 5 см/ч).
Если принять во внимание всю протяженность пути восходящего тока, то его можно будет разбить на 2 не равных по длине участка: Мертвые гистологические элементы в середине проводящего пути — сосуды или трахеиды. Протяженность значительна, но вода проходит по нему сравнительно легко, так как двигается пассивно по мертвым элементам, не испытывая с их стороны значительного сопротивления. Живые клетки корня и листа, находящиеся в начале и в конце пути передвижения. Этот путь пространственно короток, но преодолевается он с большим трудом, так как клеточные мембраны препятствуют передвижению воды. Этот ток снабжает водой все органы и ткани, приводя их в состояние тургора. Восходящий ток воды захватывает мин. ионы, поглощенные корнем, переносит их и тем самым облегчает распределение (но не поглощение!) по растению.
Концевые двигатели водного тока.
Для того чтобы вода двигалась по растению (и не просто двигалась, а поднималась вверх), необходимы затраты энергии, точки приложения которой располагаются по концам тока, вследствие чего они получили название концевых двигателей.
Нижний концевой двигатель, или корневое давление. Его роль проявляется в основном при активном поглощении — нагнетании воды. При участии актомиозиноподобных сократительных белков он не только подает воду в корневую систему, но и проталкивает ее дальше в сосуды корня и вверх по стеблю. Нагнетание воды — активный энергозависимый процесс, который сильнее всего выражен в коре корня. Сила, развиваемая концевым двигателем, невелика, она может обеспечить подъем воды на высоту не более 1 м., т. е. достаточна для травянистых растений и небольших кустарников. Верхний концевой двигатель, или присасывающая сила транспирации. При постоянном испарении воды в листьях растений разбивается сосущая сила, отсасывающая воду от ближайших клеток и передающаяся др. кл., по к-ым передвигается вода, вплоть до сосудов. В сосудах нет цитоплазмы, поэтому нет и осмотического давления, и всасывание жидкости происходит при участии всей величины сосущей силы. Она позволяет поднять воду на несколько метров. Этой силы дост. для обеспечения водой кустарников и небольших деревьев.
Поднятие воды по стволу
Многие древесные раст. имеют большую протяженность ствола. У таких растений на помощь приходят силы сцепления между молекулами воды, которые очень велики и могут достигать 30 — 35 МПа. Этой силы достаточно для того, чтобы поднять воду на 1 — 2 км, что значительно превышает высоту любого дерева. Силы сцепления молекул воды действуют только при определенных условиях: водные струи в сосудах должны идти непрерывно, без пузырьков воздуха. Если же в них попадает в-х, передвижение воды прерывается. Непрерывность водных струй в сосудах достигается их сильным натяжением и плотным прилеганием к стенкам сосудов. Водные струи оказываются как бы висящими, они удерживаются в подвешенном состоянии сосущей силой трансп. и снизу поддерживаются силой корневого Р. При сильном натяжении водных струй пр-ит натяжение и самих сосудов, при этом у последних наблюдается увелич. длины и уменьшение поперечного сечения (подобно эластичной резине). Это создает предпосылки к смыканию стенок сосудов, что привело бы к их закрыванию и прекращению водного тока. Однако этого не пр-ит, т. к. сосуды имеют внутренние утолщения стенок — кольчатые, спиральные, лестничные, к-ые наподобие каркаса растягивают их и предохр. от смыкания. Величина его незначительна — всего 20 см воды на 1 см2 площади сосудов. Однако и этого достаточно при условии не очень сильного напряжения атмосферных факторов.
Физиологическая значимость процессов минерального питания, коэффициент накопления, органогены, определение макро- и микроэлементов, их представители. Первичные и вторичные макроэлементы. Тройное правило Арнона.
Растения могут поглощать и перемещать неорганические и органические вещества. Раньше считали, что для роста растений (фотосинтезирующих автотрофов) необходимы только минеральные элементы. Однако в последние годы показано, что многие водоросли требуют витамины В12, а также веществ, содержащих кобальт и т. д.
Для наземных растений основным источником воды и минеральных веществ служит почва, не менее важна и атмосфера, которая обеспечивает фотосинтезирующие растения СО2, и, в свою очередь, сама атмосфера поглощает О2 и воду, выделяемую при транспирации.
В системе растение-почва-атмосфера происходит значительный круговорот веществ, особенно воды и СО2.
Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана с их концентрацией в среде. Например, соотношение K+/Na+ в растениях обычно составляет 5–20, тогда как в среде, как правило, больше Na+. С другой стороны, элементы, содержащиеся в окружающей среде в низких концентрациях, могут накапливаться в растениях в значительном количестве.
Эта способность растений характеризуется так называемым коэффициентом накопления:
| |
| |  |
Еще раз отметим, что ни присутствие, ни концентрация минеральных элементов в самих растениях не является критерием необходимости. Хорошо зарекомендовало себя в этом отношении (относительно необходимости) так называемое тройное правило Арнона, которое было предложено еще в 1939 году. Элемент признается необходимым в случае, когда: 1. Растение без него не может закончить свой жизненный цикл; 2. Другой элемент не может заменить функцию изучаемого элемента; 3. Элемент непосредственно включен в метаболизм растения.
Питательные элементы, которые используются растениями из субстрата в больших количествах (содержание их в золе 10–1–10–2 %), называют макроэлементами, а необходимые в значительно меньшем количестве (содержание 10–3 % и ниже) – микроэлементами.
Вода и СО2 рассматриваются отдельно, но в этом контексте необходимо считать их макроэлементами. К макроэлементам относятся K, Ca, Mg, N, P, S (Н2О и СО2), для галофитов следует добавить Na и Cl.
Микроэлементы: Mn, Zn, Cu, Cl, B, Mo, сюда же относится и Fe, который располагается на границе макро- и микроэлементов.
Водорослям необходим один или несколько из следующих элементов: Со, Si, I, V. Некоторым высшим растениям необходимы селен, кремний.
Кроме необходимых элементов для роста, в оптимальных пределах должны быть и другие факторы: рН, температура, обеспечение О2, градиент водного потенциала, а для зеленых растений – освещенность соответствующей интенсивности и протяженности.
Пассивный транспорт – перемещение веществ путем диффузии по градиенту электрохимического потенциала без затраты энергии (простая и, в какой-то мере, облегченная диффузия).
Активный транспорт – перемещение веществ против градиента электрохимического потенциала с затратой метаболической энергии, как правило в форме АТФ или редокс-цепей.
Для того чтобы понять механизмы трансмембранного переноса элементов минерального питания, остановимся на рассмотрении некоторых физико-химических закономерностях, определяющих движение ионов в растворе и мембране. Начнем с процессов пассивного транспорта (в частности вспомним законы диффузии).
Согласно первому закону Фика, поток (Ф) прямо пропорционален коэффициенту диффузии D и градиенту концентрации dC/dх в точке х в данный момент времени.
Ф = – D
Знак минус в правой части уравнения означает, что, если градиент положителен, т. е. концентрация при увеличении х возрастает, диффузионный поток направлен в противоположную сторону.
Однако при описании диффузии на большие расстояния в непрерывной системе уравнения (5.1) необходимо определить зависимость концентрации не только от расстояния, но и от времени. Поэтому целесообразно преобразовать первый закон Фика в дифференциальные уравнения в частных производных, которые обычно называют вторым законом Фика. В одномерном случае этот закон аналитически выражается следующим образом:
=
D

т. е. скорость изменения концентрации пропорциональна второй производной от концентрации на координате х. Из этого закона мы получаем очень важное соотношение:
х =
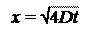
движение ионов через мембрану обусловлено градиентом электрохимического потенциала.
Обычно мембрана на внутренней стороне отрицательно заряжена по отношению к наружной поверхности. Это приводит к преимущественному поступлению катионов по сравнению с анионами. В этом случае физической движущей силой будет электрохимический потенциал  , в величину которого вносят свой вклад химический и электрический потенциалы:
, в величину которого вносят свой вклад химический и электрический потенциалы:
µi
| µi = µi + RT ln Ci + ZiFφ,
|
где – электрохимический потенциал i-иона в стандартных условиях; Zi – валентность i-иона; F – число Фарадея; φ – электрический потенциал.
Δ µi =

В реальных условиях описания процесса диффузии ионов в клетку нас интересует разность электрохимических потенциалов (Δφ) по обеим сторонам мембраны, т. е.. Используя уравнение (5.9) можно записать:

| (5.10)
|
µi = μi
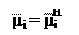
В случае равновесного состояния, когда электрохимический потенциал i-иона одинаков по обе стороны мембраны, т. е., получаем: