ФГК и является, по современным взглядам, первичным продуктом ассимиляции углерода.
Для дальнейших превращений ФГК необходимы вещества световой фазы фотосинтеза: АТФ и НАДФН. Сначала 3-ФГК фосфорилируется при участии АТФ и образуется 1,3-дифосфоглицириновая кислота. Реакция катализируется ферментом фосфоглицераткиназой.
Затем происходит восстановление за счет НАДФН и образуется фосфоглицириновый альдегид ФГА. Суммарный результат второй стадии: восстановление карбоксильной группы кислоты (–СООН) до альдегидной (–СНО). Процесс превращения катализируется дегидрогеназой фосфоглициринового альдегида. Дальнейшее превращение фосфоглициринового альдегида может происходить 4 путями.
ФГА частично с помощью триозофосфатизомеразы превращается в фосфодиоксиацетон.
Таким образом, в клетку поступают две найпростейшие формы сахаров: альдоза (ФГА) и кетоза (ФДА). Это трехуглеродные сахара (триозосахара) с присоединенной к ним фосфатной группой содержат больше химической энергии, чем ФГК. Это первые углеводы, которые образуются при фотосинтезе. С помощью альдолазы фосфодиоксиацетон (ФДА) соединяется с другой молекулой ФГА и образуется молекула фруктозо-1,6-дифосфата (ФДФ). Фруктозо-1,6-дифосфат дефосфорилируется и превращается во фруктозо-6-фосфат (Ф-6-Ф), что сопровождается накоплением в среде неорганического фосфата. Фруктозо-6-фосфат в дальнейшем может выйти из цикла и использоваться для синтеза запасных форм углеводов: сахарозы, крахмала, других полисахаридов. Однако ФГА (третий путь) может реагировать с эквимолярным количеством Ф-6-Ф, в результате образуются равные количества ксилулозо-5-фосфата и эритрозо-4-фосфата (транскетолаза). Затем эритрозо-4-фосфат реагирует с равным количеством ФДА и образуется седагептулозо-1,7-дифосфат (альдолаза), которая фосфорилируется до седагептулозо-7-фосфата с участием седагептулозодифосфатазы.
52. С2-цикл (фотодыхание), его роль в жизни растений, характеристика и локализация основных реакций.
Фотодыхание (или световое дыхание) – это индуцированное светом поглощение О2 и выделение СО2, которое наблюдается в зеленых клетках. Интенсивность фотодыхания увеличивается при повышении освещенности.
Путь восстановления СО2 при фотосинтезе М. Кальвин изучал при относительно высоких концентрациях углекислого газа. При более низких концентрациях СО2, приближающихся к атмосферным (0,045 %), в хлоропласте образуется значительное количество фосфогликолевой кислоты.
Это связано с тем, что в условиях низкого парциального давления СО2 и высокого парциального давления О2 РДФ-карбоксилаза может присоединять к рибулозо-1,5-дифосфату кислород; это значит, что она может выполнять не только карбоксилирующую, но и оксигенерирующую функции. Присоединение кислорода к молекуле рибулозодифосфата приводит к тому, что вместо двух молекул ФГК образуется одна молекула ФГК и одна молекула фосфогликолата. Последняя во время транспорта через мембрану хлоропласта дефосфорилируется при участии фосфогликолатфосфатазы и превращается в гликолат, который поступает в специальную органеллу-периксисому (рис. 2.25).
В последней содержится каталаза – фермент, разлагающий перекиси. В периксисоме гликолат вступает в реакцию с кислородом воздуха, в результате образуется глиоксилат и перекись водорода. Реакцию катализирует гликолатоксидаза.
Перекись сразу разрушается каталазой на воду и кислород, а глиоксилат в результате реакции трансаминирования, которая проходит при участии глиоксилатглицинаминотрансферазы, превращается в глицин, который перемещается в митохондрию. Тут из двух молекул глицина при участии серинглиоксилатаминотрансфераза синтезируется одна молекула серина, образуется СО2, аммиак и восстанавливается НАД+. СО2 или выделяется в атмосферу, это значит теряется для растения, или вновь может связаться в С3-цикле.
Предполагают, что образующийся НАДН может быть связан с электрон-транспортной цепью дыхания, что приводит к синтезу АТФ при его окислении.
Серин из митохондрий транспортируется вновь в периксисому, где после дезаминирования превращается в оксипируват, потом в глицерат, и наконец, в ФГК. ФГК – конечный продукт этого цикла может поступать в С3-цикл. Цикл превращения гликолата в триозофосфат называется гликолатным. Образовавшийся аммиак не выделяется в окружающую среду, а связывается в виде глутамина.
Так как образуются кислоты, содержащие 2 атома углерода, то этот цикл получил название С2-цикла. С другой стороны, так как во время этих превращений происходит поглощение кислорода и выделение СО2, то этот процесс получил название фотодыхания.
Хотя органеллой фотодыхания считается периксисома, в этом цикле непосредственное участие принимают хлоропласты и митохондрии. В клетке эти органеллы примыкают друг к другу, что также подчеркивает их функциональную зависимость. Таким образом, мы приходим к очень важному выводу: в клетке вместе с изолированностью органелл существует их интеграция.
53. С4-цикл (Хетча – Слэка – Карпилова), его роль в жизни растений, характеристика и локализация основных реакций.
Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4-пути, имеются два типа клеток и хлоропластов:1) мелкие гранальные пластиды в клетках мезофилла листа;2) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки. Клетки обкладки имеют утолщенные клеточные стенки, содержат большое количество хлоропластов и митохондрий, расположены вокруг сосудистых пучков в 1 или 2 слоя. Совокупность указанных особенностей анатомического строения получила название корончатой анатомии или корончатого синдрома (от слова kranz — корона). Хлоропласты разных типов клеток характеризуются не только особенностями строения, но и разным типам фосфорилирования. В клетках мезофилла по преимуществу происходит нециклическое фосфорилирование и образуется НАДФН, необходимый для цикла Кальвина, идущего в клетках обкладки. В хлоропластах клеток обкладки идет только циклическое фосфорилирование. Такое разделение типов фосфорилирования, возможно, связано с тем, что к хлоропластам клеток обкладки, расположенным в глубине листа, проникает по преимуществу более длинноволновый свет, который не поглощается фотосистемой, ответственной за разложение Н20. На первом этапе С4-пути углекислый газ, диффундирующий в лист через устьица, попадает в цитоплазму клеток мезофилла с мелкими хлоропластами, в которых и происходит реакция карбоксилирования фосфоенолпировиноградной кислоты (феп):
Реакция катализируется ферментом фосфоенолпируваткарбоксилазой (фепкарбоксилазой) с образованием щавелевоуксусной кислоты (оксалоацетат). ЩУК преобразуется в яблочную кислоту (малат) или аспарагиновую (аспартат). Восстановление до маната происходит в присутствии НАДФН, а для образования аспартата необходимо наличие NH4+. Затем яблочная (или аспарагиновая) кислота, по-видимому, по плазмодесмам передвигается в клетки обкладки. В клетках обкладки яблочная кислота декарбоксилируется ферментом малатдегидрогеназой до пировиноградной кислоты (пируват) и С02. Реакция декарбоксилирования может варьировать у разных групп растений с использованием разных ферментов. С02 поступает в хлоропласты клеток обкладки и включается в цикл Кальвина—присоединяется к РБФ. Пируват возвращается в клетки мезофилла и превращается в первичный акцептор С02 — ФЕП. Таким образом, при С4-пути реакция карбоксилирования происходит дважды. Это позволяет растению создавать запасы углерода в клетках. Акцепторы С02 (ФЕП и РБФ) регенерируют, что и создает возможность непрерывного функционирования циклов. Фиксация С02 с участием ФЕП и образованием малата или аспартата служит своеобразным насосом для поставки С02 в хлоропласты обкладки, функционирующих по С3-пути. Поскольку при таком механизме фотосинтеза принимают участие два типа клеток и два типа хлоропластов, этот путь называют еще кооперативным (Ю.С. Карпилов, 1970). Высказывается мнение, что С4-путь возник в процессе эволюции как приспособление к изменившимся условиям среды. При возникновении фотосинтеза атмосфера была значительно богаче С02 и беднее 02. Именно поэтому важнейший фермент цикла Кальвина Rubisco (РБФ-карбоксилаза/оксигеназа) может работать только при сравнительно высоких концентрациях С02. Благодаря деятельности самих растений состав атмосферы изменился: содержание С02 резко уменьшилось, а 02 возросло. В изменившихся условиях в осуществлении темновых реакций фотосинтеза появился ряд приспособительных черт. В частности значительно увеличилось содержание фермента Rubisco, который составляет почти половину белков стромы хлоропластов. Вместе с тем у некоторых растений выработался особый, дополнительный путь связывания С02 с помощью ФЕП-карбоксилазы. Этот фермент обладает большим сродством к углекислоте и работает при концентрациях С02 во много раз более низких по сравнению с Rubisco. Установлено, что и сопротивление мезофилла диффузии С02 у С4-растений более чем в 3,5 раз меньше и составляет 0,3—0,8 см/с, в то время как у С3 — растений — 2,8 см/с. Фиксация по С4-пути имеет еще ряд преимуществ. Растения С3-пути характеризуются высокой интенсивностью процесса, получившего название фотодыхания. Под фотодыханием понимают поглощение кислорода и выделение С02 на свету с использованием в качестве субстрата промежуточных продуктов цикла Кальвина. Как показали исследования, Rubisco (РБФ-карбоксилаза/оксигеназа) имеет двойственную функцию и может катализировать не только реакцию карбоксилирования цикла Кальвина: РБФ + С02 —> 2ФГК. Rubisco способна реагировать с 02, осуществляя оксигеназную реакцию, при этом образуется фосфогликолевая кислота: РБФ + 02 -> ФГК + фосфогликолевая кислота. Фосфогликолевая кислота через ряд превращений распадается с выделением С02. Таким образом, при фотодыхании часть промежуточных продуктов фотосинтеза теряется за счет выделения С02. Реакции окисления и карбоксилирования конкурируют друг с другом, а осуществление Rubisco карбоксилазной или оксигеназной функции зависит от содержания 02 и С02. Фотодыхание требует повышенной концентрации 02. Между тем, как уже упоминалось, в хлоропластах клеток обкладки концентрация 02 понижена, так как в них происходит только циклическое фосфорилирование, при котором вода не разлагается и 02 не выделяется. Вместе с тем в клетках обкладки повышена концентрация С02. Такие условия ингибируют процесс фотодыхания в клетках обкладки и поэтому растения С4-типа характеризуются очень низкой потерей С02 в результате фотодыхания. Потери на фотодыхание у С3-растений особенно возрастают при повышении температуры и освещенности. В этой связи понятно, что растения С4 — это, главным образом, южные и даже тропические, которые получают дополнительные преимущества в смысле продуктивности фотосинтеза. Оптимум температуры для фотосинтеза у С3-растений 20—25°С, тогда как у растений С4 30—45°С. Светонасыщение фотосинтеза С4-растений также происходит при более высоких значениях интенсивности света, чем у С3-растений. Так, у растений С3-пути интенсивность фотосинтеза перестает увеличиваться при 50% от полного солнечного освещения, в то время как у С4-форм этого не происходит. Такие особенности С4-растений объясняют высокую интенсивность фотосинтеза при повышенных температуре и освещенности. Характерным признаком растений С4-пути является, наконец, то, что образование продуктов цикла Кальвина происходит в хлоропластах, расположенных непосредственно около проводящих пучков. Это благоприятствует оттоку ассимилятов и, как следствие, повышает интенсивность фотосинтеза. Различия между С3- и С4-растениями можно продемонстрировать, поместив их рядом в одной камере (например, кукурузу и бобы) при высокой температуре и освещенности. Окажется, что С02, выделяясь в процессе дыхания, постепенно переходит к кукурузе и соответственно изменяет темпы ее роста. Кукуруза как бы «съедает» растения бобов.
54. История открытия и пространственная организация в клетках растений процессов С4-цикла.
Цикл Хетча – Слэка. Цикл Кальвина – основной, но не единственный путь восстановления СО2. Так австралийские ученые М. Хетч и К. Слэк (1966) и советский ученый Ю. Карнилов (1960) выявили, что у некоторых растений, главным образом, тропических и субтропических, таких как кукуруза, сахарный тростник, сорго и другие, основная часть меченного углерода (14СО2) уже после нескольких секунд фотосинтеза обнаруживается не в фосфоглицериновой кислоте, а в щавеливо-уксусной (ЩУК), яблочной (ЯК) и аспаргиновой (АК) кислотах. В этих кислотах можно обнаружить в первые секунды до 90 % поглощенного 14СО2. Через 5–10 минут метка появлялась в фосфоглицериновой кислоте, а затем в фосфоглицериновых сахарах. Так как эти органические кислоты содержат по 4 атома углерода, то такие растения начали называть С4-растениями в отличие от С3-растений, в которых радиоуглеродная метка появляется, прежде всего, в ФГК.
Это открытие положило начало серии исследований, в результате которых подробно был изучен химизм превращения углерода в фотосинтезе в С4-растениях. Акцептором углекислого газа в этих растениях является фосфоенолпировиноградная кислота (ФЕП).
Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4-пути, имеются два типа клеток и хлоропластов:1) мелкие гранальные пластиды в клетках мезофилла листа;2) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки. Фотосинтез растений осуществляется в хлоропластах: обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем. Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие С4 и CAM фотосинтез, сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые, соединяясь друг с другом, образуют тилакоиды, которые, в свою очередь, группируются в стопки, называемые гранами. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме. В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза. Цианобактерии, таким образом, как бы сами являются хлоропластом, и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды однако не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
55. Отличие малатного и аспартатного путей у С4-растений. Схема реакций малатного пути.
В настоящее время в зависимости от того, какая из органических кислот (малат или аспарат) декарбоксилируется, С4-растения делят на два типа: малатный тип (кукуруза, тростник) и аспаратный тип (сорго и др.).В свою очередь растения последнего типа подразделяют на две группы: растения, которые используют в реакции НАД-зависимую малатдегидрогеназу и растения, которые используют фосфоенолпируват-карбоксикиназу.Как отмечалось, С4-растения отличаются от С3-растений и по анатомии листовой пластины. Фотосинтез идет в клетках обкладки и в клетках мезофилла. Оба типа фотосинтезирующих тканей отличаются по строению хлоропластов. Хлоропласты клеток мезофилла имеют строение, присущее большинству растений: они содержат два типа тилакоидов – тилакоиды гран и тилакоиды стромы (гранальные хлоропласты). Клетки обкладки содержат более крупные хлоропласты, часто наполненные крахмальными зернами и не имеющими гран, т. е. эти хлоропласты содержат только тилакоиды стромы (агрональные).
Считают, что агрональные хлоропласты образуются в процессе онтогенеза листа из обычных гранальных хлоропластов, так как на ранних стадиях развития эти хлоропласты также имеют граны. Таким образом, для С4-растений характерны следующие особенности в строении: – многочисленные воздушные полости, по которым воздух из атмосферы подходит непосредственно к большому количеству фотосинтезирующих клеток, обеспечивая эффективное поглощение углекислоты; – слой клеток обкладки сосудистых пучков, плотно упакованных около проводящих пучков; – клетки мезофилла, которые располагаются менее плотными слоями около клеток обкладки сосудистых пучков; – большое количество плазмодесм между клетками обкладки сосудистых пучков и клетками мезофилла; – содержание как гранальных, так и агранальных хлоропластов.
Гранальные и агранальные хлоропласты отличаются и по характеру приходящих в них реакций фотосинтеза. В клетках мезофилла с мелкими гранальными хлоропластами происходит карбоксилирование ФЕП с образованием ЩУК (первичное карбоксилирование), а затем образуется малат. Малат перемещается в клетки обкладки. Тут малат окисляется и декарбоксилируется с участием малатдегидрогеназы. Образуется СО2 и пируват. СО2 используется для карбоксилирования рибулозо-1,5-дифосфата (вторичное карбоксилирование) и таким образом включается С3-цикл, который идет в агранальных хлоропластах клеток обкладки. Пируват возвращается в клетки мезофилла, где фосфорилируется за счет АТФ, что приводит к регенерации ФЕП, и цикл замыкается.
Таким образом, в С4-растениях карбоксилирование происходит два раза: в клетках мезофилла и в клетках обкладки. Что происходит в растениях, в которых из ЩУК образуется аспартат (аспарагиновая кислота)? У тех растений, которые используют для реакций декарбоксилирования НАД-зависимую малатдегидрогеназу, ЩУК трансаминируется под воздействием цитоплазматической аспартаминотрансферазы, которая использует в качестве донора глутаминовую кислоту (ГК-глутаминовая, ОГК-2-оксоглутаминовая кислота)
АК переходит из цитоплазмы клеток мезофилла в митохондрии клеток обкладки сосудистого пучка, вероятно, через плазмодесмы. Там происходит противоположная реакция трансаминирования, которая приводит к образованию ЩУК. Затем митохондриальная малатдегидрогеназа восстанавливает ЩУК до ЯК. ЯК декарбоксилируется НАД-зависимой малатдегидрогеназой с образованием ПВК и СО2.
СО2 диффундирует из митохондрий в хлоропласты, где включается в цикл Кальвина. ПВК поступает в цитоплазму, где трансаминируется аминотрансферазой; донор аминогруппы – ГК. Образовавшийся аланин (АЛ), переносится из цитоплазмы клеток обкладки в цитоплазму клеток мезофилла (вероятно, через плазмодесмы). В дальнейшем он преобразуется в ПВК (аланинаминотрансфераза); акцептором аминогруппы является ОГК (оксаглутаровая кислота). Затем ПВК переходит в хлоропласты мезофилла и превращается в ФЕП.
56. САМ-цикл, его роль в приспособлении к засушливым условиям существования. Сравнение САМ-цикла и С4-цикла.
Кислотный метаболизм толстянковых (КМТ или САМ цикл). Мы упоминали, что есть растения суккуленты, у которых устьица закрыты для уменьшения транспирации. Это особенность влияет и на фотосинтез. Эти растения поглощают большие количества углекислого газа ночью, когда устьица широко открыты. Поглощенный СО2 используется, как и у С4-растений, для карбоксилирования ФЕП. При этом образуется ЩУК (фермент ФЕП-карбоксилаза содержится в цитоплазме). ЩУК восстанавливается до малата (НАД-зависимая малатдегидрогеназа). Малат накапливается в вакуоли, поэтому ночью вакуолярный сок становится более кислым. В следующее утро малат из вакуоли транспортируется в цитоплазму. Тут происходит его окислительное декарбоксилирование при помощи НАДФ-зависимого малик энзима (малатдегидрогеназа), что приводит к образованию пирувата и СО2 и восстановлению НАДФ+. СО2 и НАДФН и используются в цикле Кальвина, который происходит в хлоропластах, а ПВК фосфорилируется с образованием ФЕП. Последний в результате ряда превращений дает ФГК, которая может также поступать в цикл Кальвина и служит источником для образования сахарозы и крахмала.
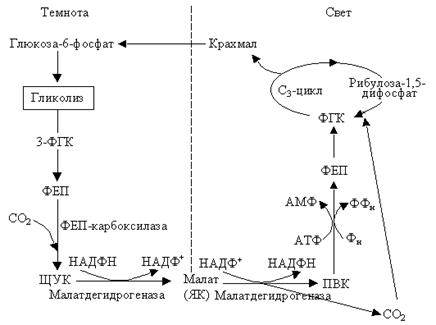
ФЕП, необходимый для акцентирования СО2, образуется ночью в процессе гликолиза. Следовательно, днем в хлоропластах идет обычный С3-цикл. По мере расходования кислот, накопленных в вакуоли, величина рН в вакуоли снова увеличивается. Для суккулентов – растений сухих мест, разделение во времени ночной фиксации СО2 и переработки СО2 на следующий день выгодно. Это позволяет им осуществлять фотосинтез при закрытых устьицах и одновременно уменьшать транспирацию. Таким образом, можно отметить, что, для САМ-растений характерны следующие особенности:
– устьица обычно открыты ночью (в темноте) и закрыты на протяжении дня. Такое состояние устьиц противоположно тому, что характерно для других растений;
– фиксация СО2 происходит в темный час суток в содержащих хлоропласты клетках фотосинтезирующих тканей листа или стебля, причем синтезируется значительное количество свободной ЯК; – ЯКкислотанакапливается в больших вакуолях, которые характерны для клеток этих растений; – на свету ЯК декарбоксилируется и освободившийся СО2 превращается за счет энергии света в сахарозу и запасной глюкан (например, крахмал) в световых реакциях фотосинтеза С3-типа. – в последующий темновой период часть запасенного глюкана разрушается с образованием молекул-акцепторов для темновой реакции фиксации СО2. Следовательно, в тканях САМ-растений существует суточный цикл: ночью содержание запасенного глюкана падает и содержание яблочной кислоты повышается, а днем происходят противоположные изменения. Следовательно, реакции разделены во времени, а у растений С4-типа процессы разделены в пространстве.
С4 и САМ-типы фиксацин СО2 являются дополнительными, облегчающими функционирование цикла Кальвина в растениях, которые приспособились к жизни в условиях повышенных температур (сахарный тростник, сорго,кукурузадр.), или недостатка влаги (суккуленты). В заключении можно дать следующую характеристику процесса фотосинтеза: – сущность фотосинтеза состоит в фотоокислении воды с выделением кислорода и использованием водорода на восстановление СО2 с образованием органических веществ; – фотосинтез состоит из двух фаз: световой и темновой; – энергия поглощенного света используется для движения электронов против градиента окислительно-восстановительного потенциала, – транспорт электронов сопряжен с синтезом АТФ; световая стадия происходит в тилакоидах стромы и гран, а темновая – в строме хлоропластов, т. е. для световой стадии необходимы мембраны.








