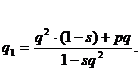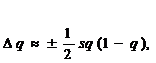(по Д. С. Фолконеру, 1985 и С. Г. Инге-Вечтомову, 1989).
| Показатели
| Генотипы
|
|
| А1А1
| А1А2
| А2А2
| всего
|
| Исходные частоты (до отбора)
| P2
| 2pq
| q2
| 1
|
| Коэффициенты отбора
| 0
| 0
| S
|
|
| Приспособленность
| 1
| 1
| 1 - s
|
|
| Вклад гамет (частоты в F 1 после отбора)
| P2
| 2pq
| q2 (1 - s)
| 1 – s q2
|
В числе прочих генотип А2А2 - это рецессивная гомозигота с коэффициентом s действующего против нее отбора. Умножение исходной частоты на приспособленность дает частоту каждого генотипа после отбора. При этом частота генотипа записывается как «вклад гамет», чтобы показать, что отбор действует в течение всего жизненного цикла. Заметим, что после отбора сумма частот уже не единица, поскольку при отборе были понесены определенные утраты, характеризуемые величиной s q 2.
Чтобы найти частоту гамет А2, а значит, и частоту гена А2 у потомков, берем вклад гамет особей А2А2 плюс половину вклада особей А1А2 и делим на новую сумму (см. уравнение 15.2).Таким образом, новая частота гена будет:
Выражение можно упростить, подставив
р = 1 - q. После преобразования получаем:
(16.4)
Изменение частоты гена D q 1, происходящее в результате одного поколения отбора, есть:
D q = q 1 - q
Подставляя для q 1 ее выражение из формулы (16.4), после некоторого преобразования получим:
(16.5)
Отсюда видно, что действие отбора на частоту гена зависит не только от интенсивности отбора
s, но также и от начальной частоты гена
q. Если и коэффициент отбора
s, и частота гена
q малы, то знаменатели стремятся к единице и ими можно пренебречь, а
D q выразить в приблизительной форме (Д. С. Фолконер, 1985). Тогда для отбора в обоих направлениях, в отсутствие доминирования, получаем:
(16.6)
a при полном доминировании:
(16.7)
Hапример, если р = 0,9, а q = 0,1, то при полном доминировании и s = 0,1,
D q = 0,1 × 0,12 × (1 - 0,1)= 0,0009,
q 1 = 0,1 - 0,0009 = 0,0991.
Скорость устранения доминантных и рецессивных аллелей из популяции различна. И доминантные и рецессивные мутации (аллели) могут быть летальными, полулетальными, определять полную или частичную стерильность, вызывать различные морфологические изменения и физиологическую (биохимическую) недостаточность. Они могут быть с полной или частичной проявляемостью (пенетрантностью) и различной экспрессивностью.
Организмы, несущие летальные доминантные гены или гены стерильности с полным проявлением, устраняются отбором в первом же поколении. Все другие доминантные гены, лишь снижающие жизнеспособность или сокращающие плодовитость, и даже летальные гены, но с не полной пенетрантностью, могут сохраняться в популяции некоторое время и лишь постепенно элиминируются в ряду поколений. Однако полной элиминации этих генов будет противодействовать процесс мутирования - повторного возникновения тех же мутаций. В случае, когда доминантные гены имеют приспособительную ценность, они подхватываются отбором, и их частота в популяции быстро возрастает. То есть, доминантный ген в каждом поколении находится под контролем отбора.
Рецессивные мутации, в отличие от доминантных, могут находиться в популяции в скрытом, гетерозиготном состоянии, накапливаться в ней, создавая огромный мутационный резерв. Чем меньше частота рецессивных аллелей в популяции, тем в большей степени гетерозиготы превосходят в численном отношении рецессивные гомозиготы. Это вытекает из формулы Харди-Вайнберга. Напомним, что число рецессивных гомозигот в популяции составляет q 2, в то время как число гетерозигот составляет 2 pq. Чем больше будут устраняться отбором из популяции гомозиготы, тем больше возрастает роль гетерозигот, которые являются поставщиками рецессивных аллелей в последующих поколениях. Следовательно, отбор рецессивных генов оказывается менее эффективным, чем доминантных.
Рассмотрим пример, иллюстрирующий это положение. Предположим, что рецессивные гомозиготы полностью устраняются, т.е. коэффициент отбора для них s =1,00. Такой случай возможен, если гомозиготы летальны или бесплодны. Допустим, что частота такого рецессивного гена в генофонде исходной популяции равна q =0,5. Согласно формуле Харди - Вайнберга, такая популяция будет состоять из 25% А1А1, 50% А1А2 и 25% А2А2.
Рассмотрим динамику изменений генотипов в ряду поколений (табл. 16.4).
Таблица 16.4
Эффективность отбора при полной элиминации рецессивных гомозигот (s =1)
|
Поколения (F)
| Частота рецессивного аллеля q
|
% особей с генотипом
|
|
|
| А2А2
| А1А2
| А1А1
|
| 1
| 0,500
| 25,00
| 50,00
| 25,00
|
| 2
| 0,333
| 11,09
| 44,42
| 44,42
|
| 3
| 0,250
| 6,25
| 37,50
| 56,25
|
| 4
| 0,200
| 4,00
| 32,00
| 64,00
|
| 5
| 0,167
| 2,78
| 27,78
| 69,44
|
| 9
| 0,100
| 1,00
| 18,00
| 81,00
|
| 10
| 0,091
| 0,83
| 16,53
| 82,64
|
| 20
| 0,048
| 0,23
| 9,07
| 90,70
|
| 30
| 0,032
| 0,10
| 6,24
| 93,65
|
| 40
| 0,024
| 0,06
| 4,76
| 95,18
|
| 50
| 0,020
| 0.04
| 3,84
| 96,12
|
| 100
| 0,010
| 0,01
| 1,96
| 98,03
|
Как показывают данные табл. 16.4 относительная частота гетерозигот в последующих поколениях будет значительно медленнее сокращаться в популяции, чем частота рецессивных гомозигот. Но и полное устранение рецессивных гомозигот в каждом поколении не приводит к окончательному их исчезновению даже в сотом поколении, так как гетерозиготные особи являются постоянными поставщиками гомозиготных рецессивов.
А если рецессив не полностью летален? Если рецессивные гены определяют различную степень понижения жизнеспособности? Тогда коэффициент отбора s будет меньше 1 (s < 1), и тогда элиминация рецессивных аллелей еще более замедляется. Например, как в таблице 16.5, когда первоначальное число гомозиготных особей равно 1%.
Таблица 16.5