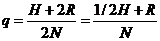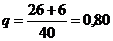Законы Менделя были выведены из данных по отдельным семьям - исследования начала века ограничивались анализом сходства и различий между родителями и их потомством. Пример: скрещивание растений с белыми и красными цветками и определение соотношения между различными функциями растений, выращенных из семьи от такого скрещивания.
Генетика популяций в отличие от посемейного анализа рассматривает статистические следствия из законов Менделя, проявляющиеся в группе семей или особей. Предмет этой науки - явление наследственности на популяционном уровне. Механизм наследственности здесь предполагается таким же, как и в менделевской генетике, но специалисты по генетике популяций изучают:
1)соотношения индивидуумов различных типов (например, растений с красными и белыми цветками);
2) частоты различных типов скрещивания;
3) соотношения различных форм, получающихся от каждого типа скрещиваний, в популяциях;
4) изменение генетической структуры популяций в поколениях при различных условиях окружающей среды (Ч. Ли, 1978).
Вернемся еще раз к термину «популяция», который мы упоминали ранее.
Популяция - это наименьшая элементарная единица эволюции. Отдельный организм - (особь) не может быть единицей эволюции, так как содержит лишь незначительную часть генетической информации вида, а с его смертью утрачивается и эта доля информации. Сам вид занимает большой ареал, содержит многообразие форм и является громоздкой системой для анализа генетических процессов, происходящих в нем. Н. В. Тимофеев – Ресовский и др. (1973), определяют популяцию как «совокупность особей определенного вида, в течение достаточно длительного времени (большого числа поколений) населяющих определенное пространство, внутри которого практически осуществляется та или иная степень панмиксии и нет заметных изоляционных барьеров, которая отделена от соседних таких же совокупностей особей данного вида той или иной степенью давления тех или иных форм изоляции».
Жизнь отдельной особи ограничена во времени, и ее генотип сохраняется постоянным в течение всей жизни, маскируя мутации. Популяция в противоположность отдельной особи практически бессмертна. Она может быть большой или малой по численности, иметь обширный или ограниченный ареал и от поколения к поколению может внезапно или постепенно изменяться по своему генетическому составу. Поэтому изучение генетики популяций неизбежно связано с изучением органической эволюции, которая с генетической точки зрения является не чем иным, как процессом кумулятивного изменения наследственных особенностей видов.
Экспериментальное изучение популяций началось с выхода в 1926 году работы С. С. Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики», которая положила начало развития в нашей стране популяционной генетики. В этой работе впервые было показано, гетерогенность является нормой для природных популяций. Здесь же было сделано три важных утверждения:
1) Мутационный процесс протекает в природных популяциях протекает точно также как и в лабораторных (например, у дрозофилы);
2) Мутации возникают у всех видов, и, как правило, новые мутации рецессивны к нормальным ("диким") аллелям;
3) Характерная черта природных популяций - преобладание панмиксии, что дает возможность описывать динамику генов с помощью закона Харди-Вайнберга.
Из экспериментальных работ С. С. Четверикова следовало, что даже в случае давления отбора, направленного на элиминацию рецессивных гомозигот, рецессивный (мутантный) ген бывает надежно укрыт от действия отбора, находясь в гетерозиготе. Мутация как бы "засосана" популяцией, но не растворена в ней. За внешней однородностью особей скрывается огромная гетерогенность.
Популяционная генетика включает как частные вопросы изучения отдельных видов, так и теоретические разработки, на базе которых можно проводить расчеты в популяциях всех видов животных, растений и человека.
Долгое время оставался дискуссионным вопрос о модели строения популяций. В 1940-50-х годах существовали две конкурирующие гипотезы о генетической структуре природных популяций:
1) классическая модель - генетическая изменчивость популяций очень мала;
2) балансовая модель - генетическая изменчивость достаточно велика.
По классической модели подавляющее число локусов содержит аллели дикого типа с частотой, близкой к единице. Кроме того, в генофонде встречаются с небольшой частотой вредные (рецессивные) мутации, не поддерживаемые отбором. Согласно этой модели, типичная особь гомозиготна по аллелям дикого типа почти по всем локусам и лишь по некоторым из них гетерозиготна. Нормальный, или идеальный, генотип гомозиготен по аллелям дикого типа. Эволюция происходит благодаря тому, что время от времени в результате мутаций появляется какой-то новый удачный аллель, частота которого постепенно возрастает под давлением естественного отбора. Таким путем новый аллель становится аллелем дикого типа, способным вытеснить старый.
Согласно балансовой модели, не существует одного аллеля дикого типа, а по каждому локусу есть серии аллелей с варьирующими частотами. Популяция состоит в основном из гетерозиготных особей. В рамках этой модели эволюция - это процесс постепенного изменения частот аллелей по многим локусам. Аллели разных локусов действуют не изолированно, а выступают в качестве элементов сложных генных систем. Влияние отдельного аллеля на приспособленность особей или популяции в целом зависит от присутствия в генотипе других аллелей, образующих единую гармоническую систему.
Изменения частот аллелей в одном локусе влекут за собой изменения частот аллелей в других локусах. Балансовая модель, так же, как и классическая, признает, что многие мутантные аллели, безусловно, вредны и элиминируются или поддерживаются естественным отбором (Ф. Айяла, 1984, цит. по С. И. Малецкий, 1995).
С течением времени явление природного полиморфизма популяций было показано на множестве объектов.
Учение о генетических процессах в популяции важно и для лесоводов, так как включает в себя вопросы динамики, численности вида, систематики и таксономии, генетический анализ.
Частота генотипов и генов
Жизнь популяции с генетической точки зрения - это динамика частот генов и генотипов в пространстве и времени.
Для того, чтобы описать генетический состав какой-либо группы особей следует определить их генотипы и выяснить, сколько особей каждого генотипа имеется в данной группе особей. Допустим, что природа фенотипических различий между генотипами нас не интересует. Предположим, что мы изучаем только два аллеля (А1 и А2), относящиеся к определенному локусу. Пусть из N диплоидных особей D несут только доминантные аллели (А1А1), Н гетерозиготные (А1А2), а R - обладают только рецессивными аллелями (А2А2), D + H + R = N. Для краткости обозначим такую группу особей следующим образом: (D, H, R). Эти три символа (D, H, R) соответствуют генотипам А1А1, А1А2, А2А2. Их соотношение описывает генетический состав популяции, а сами количественные соотношения называются частотами генотипов. Таким образом, частота данного генотипа - это его относительная численность (процент) в группе особей. Например, если обнаруживается, что пятая часть особей имеет генотип А1А1, то его частота составляет 0,20 или 20%. Естественно, что частоты всех имеющихся генотипов в сумме должны составлять единицу или 100%.
Поскольку популяция - это не просто механическая смесь особей, а размножающаяся группа, то популяционная генетика изучает не только генетический состав этой группы, но и динамику ее в ряду поколений. В каждом следующем поколении из передаваемых с гаметами родительских генов у потомков составляется новый набор генотипов. Преемственность поколений поддерживается в первую очередь генами, которые несет данная популяция, а не генотипами, поскольку последние расщепляются (Д. С. Фолконер, 1985). Поэтому генетический состав популяции описывается множеством генных частот, которые представляют собой соотношение аллелей каждого локуса. Если, например, А1 - аллель в локусе А, то частота гена А1 есть относительное число, или процент, всех генов этого локуса, находящихся в аллельном состоянии А1. Для любого локуса частоты его аллелей в сумме должны составлять единицу, или 100%.
|
| | 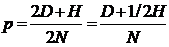 |
Частоты генов данного локуса в группе особей можно определить, зная частоты генотипов. Предположим, что наличие особей трех типов в исследуемой группе (
D, H, R) обусловлено существованием только двух аллельных генов
А1 и
А2 , и в сумме
N особей несут
2 N генов. Поскольку каждая особь
А1А1 имеет два гена
А1, а каждая особь
А1А2 - лишь один ген этого типа, полное число генов
А1 в группе составляет
2 D + H. Следовательно, доля генов
А1 равна:
(15.1)
Это отношение называется частотой гена А1
Аналогично определяется частота гена А2:.
(15.2)
Так что p + q = 1.
Например, в группе из 40 особей (2, 12, 26)
Нередко фактические численности особей этих трех типов заменяют их долями от общего числа особей. Такая форма замены особенно удобна, когда имеют дело с большой по численности группой. В этом случае принимают D + H + R =1, где D означает теперь долю особей А1А1 в группе, H - долю особей А1А2 и R - долю особей А2А2. Отсюда следует, что
p = D + 1/2 H (15.1¢)
и
q = 1/2 H + R (15.2¢)
В таких обозначениях приведенную выше популяцию (2, 12, 26) можно представить в виде (0,05, 0,30, 0,65), где, как и прежде p = 0,05 + 0,15 = 0,20 и q = 0,15 + 0,65 = 0,80.
При исследовании лесных древесных пород, как правило, рассматривают именно большие популяции и символы D, H, R будут обозначать доли (частоты) трех названных генотипов.
Закон Харди-Вайнберга
Этот закон гласит, что если популяция находится в равновесии, то частота генотипов и генов у родителей и потомства не меняется. Рассмотрим большую панмиктическую популяцию, в которой аллель А1 встречается с частотой р, а аллель А2 с частотой q, при этом p + q = 1. Если частоты трех генотипов, соответствующих данной паре аллелей, равны:
А1А1 = D = p 2, А1А2 = H = 2 pq, А2А2 = R = q 2,
то их пропорции в следующем поколении останутся такими же, как и в предыдущем. Заметим также и здесь, что p 2 + 2 pq + q 2 = 1. В этом случае говорят, что популяция (p 2, 2 pq, q 2) находится в равновесии при условии случайного скрещивания. Под «равновесием» понимается отсутствие изменений частот генотипов в последовательных поколениях одной популяции. Очевидно, что в этих условиях и частоты обоих генов также остаются неизменными.
Особое состояние равновесия, обусловленное случайным скрещиванием, описывается с помощью закона или правила Харди-Вайнберга (английский математик Дж. Харди и немецкий биолог Г. Вайнберг установили его независимо друг от друга в одном и том же 1908 г.). Для генетики популяций этот закон имеет фундаментальное значение, поэтому мы рассмотрим его более подробно. Если скрещивание осуществляется случайным образом, а популяция достаточно велика, частоты различных типов скрещивания можно определить, заменив D на р2 и т.д., как это показано в табл. 15.1.
Cледует, однако, подчеркнуть, что частоты, полученные теоретически, будут реализовываться только в очень больших по численности популяциях. Как синоним случайного скрещивания иногда используют термин «панмиксия» и в этом случае говорят, что популяция является панмиктической.
Если объединить реципрокные скрещивания (это допустимо, поскольку мы рассматриваем аутосомные гены), то девять различных вариантов скрещивания можно подразделить на шесть типов.
Таблица 15.1