

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Археология об основании Рима: Новые раскопки проясняют и такой острый дискуссионный вопрос, как дата самого возникновения Рима...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Археология об основании Рима: Новые раскопки проясняют и такой острый дискуссионный вопрос, как дата самого возникновения Рима...
Топ:
Определение места расположения распределительного центра: Фирма реализует продукцию на рынках сбыта и имеет постоянных поставщиков в разных регионах. Увеличение объема продаж...
Интересное:
Принципы управления денежными потоками: одним из методов контроля за состоянием денежной наличности является...
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Искусственное повышение поверхности территории: Варианты искусственного повышения поверхности территории необходимо выбирать на основе анализа следующих характеристик защищаемой территории...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
План лекции:
1. Системы классификации и таксономия дрожжей
2. Строение дрожжевой клетки.
3. Питание и метаболизм дрожжей.
4. Генетика дрожжей.
5. Микробиологические аспекты практического использования дрожжей.
СИСТЕМЫ КЛАССИФИКАЦИИ И ТАКСОНОМИЯ ДРОЖЖЕЙ
Хотя характерная почкующаяся форма дрожжей была зафиксирована еще ван Левенгуком в 1680 г., более детальное описание и идентификация дрожжей продолжали оставаться сложной задачей. Поскольку у вегетативных форм большинства дрожжей нет каких-либо характерных морфологических особенностей, их нелегко идентифицировать путем визуального наблюдения. Первоначально название Saccharomyces употреблялось по отношению ко всем дрожжам, выделенным из спиртных напитков, и Мейен в 1837 г. различал в соответствии с их источником три вида Saccharomyces: S. vini — из вина, S. cerevisiae — из пива и 5. ротогит — из сидра. Половые споры у дрожжей были обнаружены в 1837 г. Шванном, но только в 1870 г. к роду Saccharomyces стали причислять исключительно такие дрожжи, которые образуют споры.
Род Saccharomyces включает около 40 видов; все они образуют путем почкования сферические или эллипсоидные клетки, производят аскоспоры в асках и обладают способностью эффективно превращать сахара в спирт. Наиболее важный в пивоварении вид Saccharomyces был выделен в чистой культуре и описан Хансеном под названием Saccharomyces cerevisiae var. ellipsoideus (Hansen) Dekker. Штаммы, отнесенные к S. cerevisiae, получили широкое распространение в пивоварении, производстве спирта, приготовлении вина, а также в получении пекарских дрожжей и биомассы. Однако штаммы дрожжей в разное время классифицировали по-разному. Барнетт, Пейн и Ярроу в своей монографии, опубликованной в 1980 г., включили в S. cerevisiae штаммы, которые по предыдущей классификации относились к восемнадцати различным видам. Один из этих штаммов — S. carlsbergensis — представляет особый интерес, так как это «дрожжи низового брожения», применяемые для приготовления легкого пива. Их отличает способность усваивать сахар мелибиозу. Этот штамм, известный в пивоваренной промышленности как S. carlsbergensis, был отнесен в 1970 г. Ван де Вальтом к S. uvarum, апозднее включен Барнеттом, Пейном и Ярроу в вид S. cerevisiae.
СТРОЕНИЕ ДРОЖЖЕВОЙ КЛЕТКИ
Клетки Saccharomyces cerevisiae имеют округлую, яйцевидную или эллипсоидную форму; размер их колеблется от 2,5 до 10 мкм в поперечнике и от 4,5 до 21 мкм в длину. Характерные особенности типичной дрожжевой клетки показаны на рис. 2-1. Можно видеть, что клетка окружена толстой клеточной стенкой. Внутри различимы многие органеллы типичной клетки: плазмалемма, ядро, митохондрии, эндоплазматический ретикулум, вакуоли, пузырьки и гранулы. Структура и функция этих клеточных органелл будут рассмотрены ниже.
Отличительная особенность популяции растущих дрожжевых клеток — наличие почек, образующихся при делении клеток. Дочерняя клетка возникает в виде маленькой почки, которая растет в течение большей части клеточного цикла, пока не достигнет размера материнской клетки. Рост дрожжей происходит в основном во время формирования почек, поэтому почка к моменту ее отделения становится по размеру более или менее такой же, как зрелая клетка. Клетки могут разойтись вскоре после деления, однако часто еще до их расхождения начинаются новые циклы клеточного деления, в результате чего образуются группы клеток. На месте отделения клеток друг от друга остаются следы, называемые у материнской клетки дочерним шрамом, а у дочерней клетки — родовым шрамом.

Дочерние и родовые шрамы можно разглядеть с помощью флуоресцентной микроскопии, а также в сканирующем электронном микроскопе. На одном и том же месте клеточной стенки Saccharomyces cerevisiae никогда не появляются две почки, таким образом, каждый раз почка оставляет новый дочерний шрам на стенке материнской клетки. Подсчитав число таких шрамов, можно определить, сколько почек образовала данная клетка. По этой величине можно оценивать возраст клетки.
КЛЕТОЧНАЯ СТЕНКА представляет собой жесткую структуру толщиной 25 нм, составляющую около 25% сухого веса клетки и состоит в основном из глюкана и маннана; наряду с этими компонентами в стенке присутствуют хитин и белок. Глюкан — это сложный разветвленный полимер глюкозы, располагающийся во внутреннем слое клеточной стенки, прилежащем к плазмалемме. По-видимому, глюкан — основной структурный компонент клеточной стенки, поскольку при его удалении она полностью разрушается. Маннан (сложный полимер маннозы) находится главным образом во внешних слоях клеточной стенки. Третий углеводный компонент стенки, хитин, представляет собой полимер N-ацетилглюкозамина; он обнаруживается в участках клеточной стенки S. cerevisiae, ассоциированных с дочерними шрамами, образуя кольцо вокруг них. Белок составляет 10% сухого веса клеточной стенки. По крайней мере часть этого белка находится в виде связанных со стенкой ферментов. У дрожжей описано несколько таких ферментов, в том числе инвертаза, щелочная фосфатаза и липаза, а также глюканаза и маннаназа— два фермента, которые, видимо, участвуют в размягчении клеточной стенки, способствуя формированию почек.
Детальная организация клеточной стенки еще не совсем ясна, однако современные теории отдают предпочтение модели трехслойной структуры, согласно которой внутренний глюкановый слой отделен от внешнего маннанового промежуточным слоем с повышенным содержанием белка (рис. 2-2, А).
КЛЕТОЧНАЯ МЕМБРАНА, или плазмалемма, дрожжевой клетки выглядит под электронным микроскопом как трехслойная структура, тесно прилегающая к внутренней поверхности клеточной стенки. Поверхность плазмалеммы обычно ровная, но на определенных этапах роста клетки на ней можно заметить впячивания.
Плазмалемма состоит примерно из равного количества липидов и белков и из небольшого количества углеводов. Лилиды представлены в основном моно-, ди- и триглицеридами, глицерофосфатидами и стеролами — эргостеролом и зимостеролом. Природа белков плазмалеммы известна хуже, но в их состав, вероятно, входят ферменты, принимающие участие в усвоении сахаров и аминокислот.
На рис. 2-2, Б показана модель структуры плазмалеммы. Каждая молекула фосфолипида состоит из двух областей: гидрофобной, т. е. отталкивающей воду, и

гидрофильной, т. е. притягивающей воду. Считается, что эти области расположены таким образом, что гидрофильные части молекулы находятся на внешней стороне мембраны, а гидрофобные — на внутренней. Молекулы белка, возможно, размещаются на поверхности мембраны или проникают внутрь ее.
Плазмалемма — важная органелла клетки. Она выполняет роль барьера проницаемости вокруг содержимого клетки и контролирует транспорт растворенных веществ внутрь клетки и из нее. Твердо доказано, что плазмалемма участвует также в регуляции биосинтеза клеточной стенки у дрожжей.
Ядро обычно расположено между вакуолью и почкой. В ядре можно различить хроматиновые тельца. Ядерная мембрана остается интактной на протяжении всего клеточного цикла. Под электронным микроскопом она выглядит как двойная мембрана, усеянная порами.
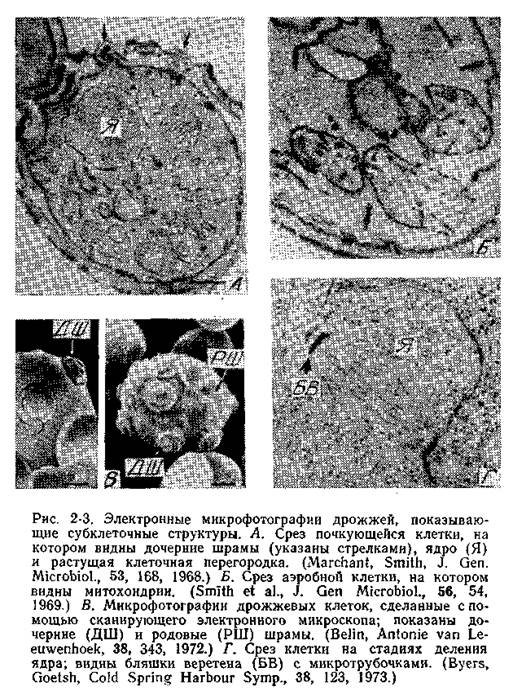
МИТОХОНДРИИ легко различимы как сферические или палочко-видные структуры, окруженные двойной мембраной. Складки внутренней мембраны образуют кристы (рис. 2-3, Б). Изучению структуры митохондрий и анализу распределения митохондриальных ферментов между мембраной и матриксом митохондрии посвящено множество работ. Большая часть ферментов цикла трикарбоновых кислот находится в матриксе митохондрии, в то время как ферменты, участвующие в транспорте электронов и окислительном фосфорилировании, локализованы на внутренней мембране митохондрии, в том числе и на кристах.
Цитоплазма дрожжевой клетки содержит систему двойных мембран, известную как эндоплазматический ретикулум. Часть этих мембран связана с рибосома-ми, как и в других организмах; однако эндоплазматический ретикулум, видимо, принимает участие и во многих иных клеточных функциях. Эндоплазматический ретикулум участвует также в образовании пузырьков, находящихся в клетке. Этим он напоминает аппарат Гольджи некоторых других организмов.
В цитоплазме встречаются и липидные гранулы, которые происходят, вероятно, также от андоплазматического ретикулума.
Зрелые дрожжевые клетки содержат большую вакуоль. В тот момент клеточного цикла, когда начинается образование почки, вакуоль, видимо, дробится на более мелкие вакуоли, которые распределяются между материнской клеткой и почкой. В дальнейшем эти маленькие вакуоли снова сливаются, образуя в материнской и дочерней клетках по одной вакуоли.
Функция вакуоли точно не установлена. Показано, что в ней содержатся гидролитические ферменты, полифосфаты, липиды, низкомолекулярные клеточные интермедиаты и ионы металлов. Вакуоль, возможно, действует как резервуар для хранения питательных веществ и гидролитических ферментов.
|
|
|

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
© cyberpedia.su 2017-2026 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

