Впервые анаэробные серовосстанавливающие экстремально-термофильные архебактерии были выделены Штеттером и Циллигом в 1981 г. из серосодержащих грязей со дна океана и глубоководных гидротермальных источников, расположенных вблизи потухших вулканов в Италии и Исландии. Открытые бактерии были отнесены к порядку Thermoproteales и разделены на семейства - Thermoproteaceae, Desulfurococcaceae, Thermococcaceae. Четвертое семейство пока не получило официального наименования, но условно названо "Stiff filaments"- "упругие нити".
Таксономическое положение семейств порядка Thermoproteales было определено на основании данных по перекрестной гибридизации 16S рРНК и ДНК различных архебактерий. Дендрограмма, полученная на основании этих результатов (рисунок 1), в основном совпадает с дендрограммой, представленной Вузом. Согласно проведенным экспериментам представители порядка Thermoprotealee попадают в одну подгруппу с Sulfolobus и вместе с Sulfolobus образуют отдельную ветвь архебактерий.

Рисунок 1. Дендрограмма функциональной стабильности гибридомов между 16S РНК и ДНК архебактерий.
В настоящее время группа анаэробных серовосстанавливающих бактерий насчитывает 7 родов (таблица 3). В состав группы входят бактерии с разной морфологией (таблица 4). Среди них есть неподвижные и подвижные формы, передвигающиеся с помощью полярно расположенных жгутиков.
Размножение отдельных видов происходит почкованием (Thermoproteus tenax, Thermophilum pendens). Почка образуется главным образом терминально, но также может образовываться на различных частях клеточной поверхности.
Таблица 3.- Анаэробные серовосстанавливающие архебактерии
| Микроорганизмы
| Год открытия
| Место
обитания
| Оптимальная температура
| Оптимальный
рН
| Содержание ГЦ в ДНК, мол. %
|
| Thermoproteus
tenax
|
| морская донная грязь в
Исландии
|
| 5,5
| 55,5
|
| Desulfurococcus D.mobilis D.mucosus
|
| кислые горячие источники в Исландии
|
| 5,5-6,0
| 50,8
51,3
|
| Thermophilum pendens
|
| серные горячие источники в Исландии
| 88-90
| 5,0-6,0
| 57,4
|
| Thermococcus celer
|
| горячие источники вулканического происхождения в Италии
|
| 5,8
| нд*
|
| Thermodiscus
maritimus
|
| горячие
источники вулканического происхождения в Италии
|
| 6,5
| нд *
|
| Pyrodictium P.occultum P.brockli
|
| донные морские гидротермы в Италии
|
|
6,5
6,5
|
51,6-56,6
|
| Pyrococcus
furiosus
|
| геотермально обогреваемые морские осадки в Италии
|
| 7,0
| нд*
|
| *нд - нет данных
|
У некоторых представителей анаэробных серовосстанавливающих
архебактерий обнаружены пили. У Pyrodictium найдена необычная сеть
фибрилл, которая часто бывает представлена в виде связок. Высказывается мнение, что сеть фибрилл предохраняет клетки от сверхвысокой температуры и принимает участие в метаболизме.
В качестве запасного вещества продуцируют гликоген (Thermococcus
Desulfurococcus, Thermoproteus).
Клеточная стенка серовосстанавливающих архебактерий построена из гликопротеиновых субъединиц гексагонального строения. Исключение составляет Pyrodictium brockii. Преобладающим компонентом клеточной стенки этого организма является белок с молекулярной массой 150000. Следует отметить, что субъединицы гексагонального строения у анаэробных бактерий встречаются редко и известны только у экстремально-термофильных бактерий. Предполагается, что гексагональное расположение субъединиц в поверхностном слое клетки обеспечивает более хорошее покрытие нижних слоев.
Различные виды анаэробных серовосстанавливающих архебактерий имеют температурный оптимум для роста в области 85-I05°C (таблица 3). Максимальная температура их роста находится в пределах 90-110°С. Среди них встречаются ацидофилы и нейтрофилы (таблица 3). Некоторые виды (Pyrodictium) устойчивы к высоким концентрациям NaCL (12 %).
В группе анаэробных серовосстанавливающих бактерий известны облигатные и факультативные автотрофы, гетеротрофы (таблица 4). Механизм фиксации СО2 у автотрофных Thermoproteales пока на расшифрован. Из органических соединений используют пептиды, аминокислоты (Desulfurococcus, Thermococcus), углеводы, например, крахмал, мальтозу (Pyrococcus), гликоген, глюкозу (Thermoproteus), а также простые соединения, такие, как формальдегид, формиат, малат, фумарат, метанол, этанол (Thermoproteus).
Распространенной формой энергетического метаболизма серовосстанавливающих архебактерий является "анаэробное дыхание". Конечным акцептором электронов служит молекулярная сера, которая восстанавливается до H2S. Из неорганических доноров электронов и энергии ряд представителей Thermoproteales (Thermoproteus, Thermodiscus, Pyrodictium) могут использовать Н2. Представители двух родов этого порядка – Desulfurococcus и Thermococcus – способны к брожению.
Таблица 4.- Характеристика некоторых свойств анаэробных серовосстанавливающих архебактерий
| Микроорганизмы
| Морфология
| Подвижность
| Способ питания
|
| Thermoproteus
tenax
| Палочки, весьма вариабельны 0,5x1-80 мкм. Иногда дают разветвления без септ. Бывают V-образной формы
| неподвижны
| факультативные автотрофы
|
| Desulfurococcus mobilis
| Кокки, вариабельны 0,2-5 мкм
| подвижны, жгутики монополярные политрихи
| гетеротроф
|
| Thermophilum pendens
| Нитевидные палочки: 0,17-0,35х1-100 мкм. Иногда дают разветвления
| неподвижны
| гетеротроф
|
| Thermococcus celer
| Клетки сферические I мкм
| подвижны, жгутики монополярные политрихи
| гетеротроф
|
| Thermodiscus maritimus
| Клетки диcковидные
| нд*
| облигатный
автотроф
|
| Pyrodictium
occultum
| Клетки в форме плоских дисков 0,2x0,3-2,5 мкм
| нд*
| облигатный
автотроф
|
| Pyrococcus
furiosus
| Клетки сферические 0,8-2,5 мкм
| подвижны, жгутики монополярные политрихи
| гетеротроф
|
| *нд - нет данных
|
В природной обстановке серовосстанавливащие анаэробные архебактерии находятся в тесном сообществе друг с другом и некоторыми термофильными архебактериями. Предполагают, что распространенные в гидротермальных источниках хемолитотрофные бактерии Thermodiscus и Thermoproteus наряду с Sulfolobus и термофильными метаногенами являются продуцентами органического вещества, которое используется гетеротрофными организмами Thermophilum и Desulfurococcus.
Галофильные архебактерии.
Группа экстремально галофильных архебактерий включает микроорганизмы, которые обитают в местах с высоким содержанием Nа, СL, Мg т.д. Лучше всего они растут при концентрации NaCl от 3 - 4,5 М, но могут расти и в насыщенном растворе NаСL (5,2 М). Высоки потребности галофилов и в Мg++- 0,1-0,5 М, а для некоторых видов (Halobacterium sodomens) - 0,6-1,2 М. Потребность в К+ значительно ниже - 0,025 М. Однако внутриклеточная концентрация К+ в клетках всегда выше (более 3 М), чем Nа+ и Мg++, и может составлять у отдельных микроорганизмов от 30 до 40 % сухого вещества клеток. Путем поддержания повышенной концентрации К+ в цитоплазме экстремальные галофилы компенсируют высокое осмотическое давление среды. Высокие концентрации ионов необходимы галофилам и по ряду других причин. Во-первых, для обеспечения структуры и жесткости клетки (Halobacterium). Во-вторых, для эффективного функционирования белоксинтезирующей и транспортной систем. В-третьих, для поддержания каталитической активности и стабильности ферментов.
В состав группы экстремальных галофилов входят морфологически разнообразные формы (таблица 5), подвижные (Halobacterium, "квадратная бактерия") и неподвижные. Размножение происходит бинарным делением в одной (Halobacterium, Halococcus) или в 2 плоскостях ("квадратная бактерия"). У некоторых видов обнаружены газовые вакуоли. В качестве запасного вещества синтезируют поли-β-гидрооксибурат. Ряд видов продуцирует галоцин Н4 (бактериоцин), подавляющий рост других галобактерий. Бактериоцидное действие галоцина Н4 связано с нарушением ионной проницаемости мембраны.
Клеточные стенки галофильных архебактерий образованы либо белковыми субъединицами, либо гликопротеиновыми, либо высокосульфатированными гетерополисахаридами.
Таблица 5.- Экстремально галофильные архебактерии
| Микроорганизмы
| Год открытия
| Морфология
|
| Halococcus
|
| кокки 0,6-1,5 мкм
|
| Halobacterium
|
| палочки 0,6-1x1-6 мкм; редко дискообразные
|
| "Квадратная бактерия"*
|
| клетки в форме квадрата; 1,5x1,5 до 11x11 мкм.
|
| Haloarcula
|
| клетки плоские, прямоугольные, в форме коробочки
|
| Natronococcus
|
| кокки
|
| Natronobacteriua
|
| палочки
|
| "Треугольная бактерия"*
|
| клетки плоские, треугольные
|
| *- название рода еще не предложено
|
Мембранный аппарат представлен цитоплазматической мембраной. Липидный состав весьма своеобразен. Основную массу составляют полярные липиды, построенные из глицерина и С20-спирта (рисунок 2). Нейтральные липиды представлены изопреновыми углеводородами. Последние, и среди них бактериоруберин, защищают клетки от фотохимических повреждений и обусловливают окраску колонии от розового до красного.
В мембране галофильных бактерий локализованы 3 небольших ре-тинальсодержащих белка, сходных с родопсином - зрительным пигментом сетчатки позвоночных. Основная доля приходится на бактериородопсин. Он содержится в пурпурных мембранах и функционирует как "протонный насос", используя энергию света. Второй ретинальсодержащий белок - галородопсин - также принимает участие в процессе утилизации энергии света. Действует как "хлоридная помпа" - закачивает на свету в клетки СL. Предполагаемый физиологический смысл этого процесса состоит в восполнении клетками потерь сL-, которые происходят под действием трансмембранного градиента электрического потенциала. Третий сенсорный пигмент - "медленный родопсин" - выполняет роль рецептора при фототаксисе (положительном и отрицательном) и обеспечивает пространственную ориентацию клеток, оптимальную для жизнедеятельности.
Галофильные архебактерии являются аэробными и факультативно анаэробными организмами. Хорошо растут при температуре 30-50°С, с оптимумом З5-40°С. Нейтрофилы. Пределы роста при рН 5,8-8,0.
Галофильные архебактерии имеют сложные пищевые потребности. Основным источником углерода и энергии служат аминокислоты и пептиды. Ряд галофилов способен утилизировать углеводы (с образованием кислоты), дикарбоновые кислоты, спирты, крахмал.
Распространенной формой энергетического метаболизма у галофильных архебактерии является аэробное дыхание. Некоторые виды в анаэробных условиях осуществляют "нитратное дыхание" и брожение. При отсутствии кислорода галобактерии могут получать всю необходимую им энергию за счет уникального процесса фотосинтеза, в котором не участвуют хлорофилсодержащие реакционные центры, характерные для фотосинтезирующих организмов. В основе фотосинтеза бесхлорофильного типа галофильных архебактерий лежат светозависимые циклические превращения бакгериородопсина в ряд промежуточных форм, которые участвуют в переносе протона через мембрану в окружающую среду. В результате между внутренней и наружной сторонами мембраны создается электрохимический трансмембранный градиент протонов. Поскольку при этом концентрация ионов Н+ снаружи выше, чем внутри, эти ионы стремятся диффундировать обратно в клетку через находящиеся в мембране молекулы АТФ-синтезирующего фермента. Проходя через протонзависимую АТФ-азу, ионы Н+ отдают свою энергию, которая используется для синтеза АТФ из АДФ и фосфора (рисунок 2).
За счет энергии трансмембранного градиента протонов поддерживается градиент Na+ между окружающей средой и цитоплазмой (антипорт H+/Na+).
Естественной экологической нишей экстремально галофильных архебактерий являются природные соленые водоемы, где в подходящих
концентрациях имеется NaCl и другие необходимые им ионы. Галофилы заселяют бассейны для выпаривания соли (т.е. солеварни) под действием солнечного света, а также белковые материалы, сохраняемые с применением соли.
Место галофилов в микробной экосистеме связано с участием в циклах углерода и азота в засоленных нишах.
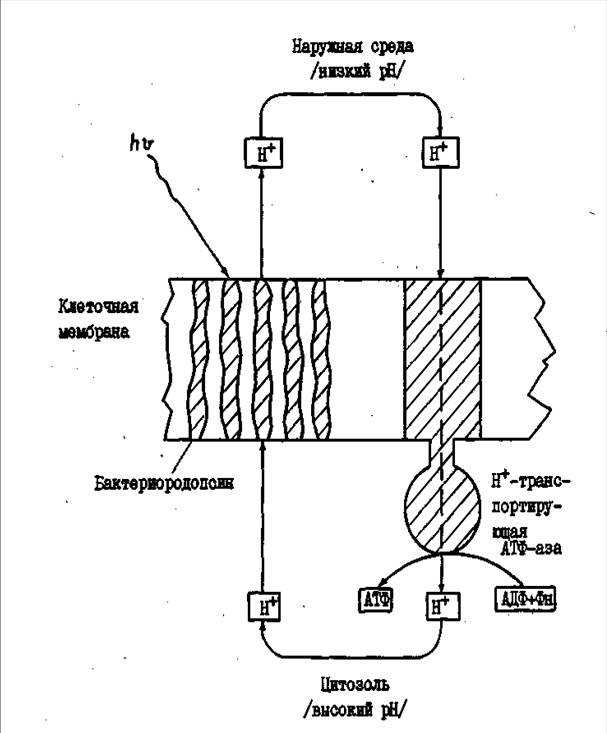
Рисунок 2. Схематическое изображение бактериородопсиновой протонной помпы








