Метанотрофы по способу питания относят к автотрофныммикроорганизмам. Это проявляется как в способности метанотрофов синтезировать все вещества клетки из С1-соединений, так и функционированию развитого у них механизма ассимиляции СО2 в восстановительном пентозофосфатном цикле.
Помимо метана в качестве единственного источника углерода и энергии облигатные метанотрофы могут использовать метанол, формальдегид и другие С1-соединения, а факультативные — же С2-, С4-кислоты, этанол, глюкозу. Использование метанотрофами С1-соединений в конструктивном и энергетическом метаболизме привело к формированию у них специфических путей ассимиляции и диссимиляции этих соединений.
Процесс полного окисления метана может быть представлен в виде следующей схемы (рисунок 1).
Первый этап — окисление метана до метанола катализируется метанмонооксигеназой в реакции:
СН4 + О2 + Н2А— СН3ОН + А + Н2О,
где О2 — молекулярный кислород, а Н2А — восстановитель. Из приведенной реакции становится понятной облигатная зависимость окисления метана от молекулярного кислорода.
Описаны две формы метанмонооксигеназы: связанная с внутрицитоплазматическими мембранами и растворимая. Остальные этапы окисления катализируются соответствующими дегидрогеназами, различающимися строением, природой акцепторов электронов и другими параметрами.
Формальдегид у метилотрофов является ключевым метаболитом, на уровне которого расходятся конструктивные и энергетические пути. Часть формальдегида превращается в вещества клетки по специфическим для этих эубактерий ассимиляционным циклическим путям, большая часть окисляется до СО2 в линейной последовательности реакций через формиат.
Дыхательные цепи метилотрофов по составу переносчиков и их локализации на мембране похожи на таковые большинства аэробных эубактерий, что предполагает возможность функционирования у них трех пунктов сопряжения. В окислительном метаболизме С1-соединений участвуют НАД, флавины, хиноны, цитохромы.
Этап окисления метана до метанола, катализируемый оксигеназным ферментом, не связан с получением клеткой энергии. Энергетическая эффективность окисления С1-соединений соответствующими дегидрогеназами определяется местом поступления электронов в дыхательную цепь. Окисление метанола до формальдегида, катализируемое ферментом, содержащим в качестве простетической группы особый хинон, сопровождается передачей электронов в дыхательную цепь на уровне цитохрома с. Это приводит к синтезу одной молекулы АТФ, т. е. указывает на функционирование только третьего пункта сопряжения (см. рисунок 1).
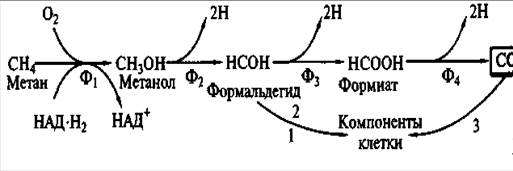
Рисунок. 1.- Окисление метана и связь энергетического и конструктивного метаболизма у метилотрофов
Ф1, — метанмонооксигеназа; Ф2 — метанолдегидрогеназа; Ф3 —формальдегиддегидрогеназа; Ф4 — формиатдегидрогеназа; ассимиляционные циклы:
1 — риболузомонофосфатный; 2 — сериновый; 3 — восстановительный пентозофосфатный.
Окисление формальдегида и формиата, зависимое от НАД, позволяет предполагать, что перенос пары электронов может быть связан с тремя трансмембранными перемещениями протонов. Полученные экспериментальные данные указывают, однако, на меньшие выходы АТФ.
У метилотрофов функционируют циклические пути ассимиляции С1-соединений, ответственные за превращения их в вещества клетки: восстановительный пентозофосфатный, рибулозомонофосфатный и сериновый. В восстановительном пентозофосфатном цикле происходит ассимиляция СО2, образующейся при окислении С1 -соединений.
В рибулозомонофосфатном и сериновом циклах обеспечивается использование в биосинтетических процессах формальдегида, образуемого при окислении разных С1-соединений. Эти реакции аналогичны таковым восстановительного пентозофосфатного цикла. Три оборота рибулозомонофосфатного цикла приводят к синтезу молекулы фосфодиоксиацетона, используемого в биосинтетических процессах.
Сериновый цикл существенно отличается от предыдущего пути ассимиляции формальдегида природой интермедиатов и ферментами. Эта реакция примечательна тем, что в сериновый цикл вовлекается СО2. Последующая серия реакций приводит к регенерированию глицина, и цикл замыкается.
Ассимиляция формальдегида через рибулозомонофосфатный цикл характерна для метилотрофов, имеющих мембранную организацию I типа, а через сериновый — для метилотрофов с системой внутрицитоплазматических мембран II типа.
ЦТК в системе катаболических путей не занимает ведущего места. У ряда облигатных метилотрофов он не «замкнут». Если даже содержит все ферменты, активность некоторых из них невысока.








