

Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Топ:
Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов...
Выпускная квалификационная работа: Основная часть ВКР, как правило, состоит из двух-трех глав, каждая из которых, в свою очередь...
Интересное:
Мероприятия для защиты от морозного пучения грунтов: Инженерная защита от морозного (криогенного) пучения грунтов необходима для легких малоэтажных зданий и других сооружений...
Искусственное повышение поверхности территории: Варианты искусственного повышения поверхности территории необходимо выбирать на основе анализа следующих характеристик защищаемой территории...
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
Известно 5 групп эубактерий, способных преобразовывать световую энергию в химическую с помощью хлорофилла. Фотосинтез, осуществляемый ими, делится на 2 типа: не сопровождающийся выделением молекулярного кислорода (бескислородный фотосинтез) и сопровождающийся выделением О2 (кислородный фотосинтез). В соответствии с этим все фотосинтезирующие эубактерий в IX издании Определителя бактерий Берджи предложено разделить на две таксономические группы в ранге классов: Апохуphotobacteria и Oxyphotobacteria. Эубактерий, осуществляющие бескислородный фотосинтез, на основании таких признаков, как пигментный состав и тонкое строение фотосинтетического аппарата, делятся на 3 группы: пурпурные, зеленые бактерии и гелиобактерии. Эубактерий, фотосинтез которых сопровождается выделением О2, включают 2 группы организмов: цианобактерии и прохлорофиты.
Цианобактерии
К цианобактериям относится большая группа организмов, сочетающих прокариотное строение клетки со способностью осуществлять фотосинтез, сопровождающийся выделением О2, что свойственно разным группам водорослей и высших растений. Объединение черт, присущих организмам, относящимся к разным царствам или даже надцарствам живой природы, сделало цианобактерии объектом борьбы за принадлежность к низшим растениям (водорослям) или бактериям (прокариотам).
На основании изучение цитологии клеток синезеленых водорослей с помощью современных методов последние отнесены к типичными прокариотам. Как следствие этого Р.Стейниером было предложено отказаться от названия «синезеленые водоросли» и называть данные организмы «цианобактериями» — термином, отражающим их истинную биологическую природу. Воссоединение цианобактерий с остальными прокариотами поставило исследователей перед необходимостью пересмотра существующей классификации этих организмов и подчинения ее правилам Международного кодекса номенклатуры бактерий.
|
|
Цианобактерии — морфологически разнообразная группа грамотрицательных эубактерий, включающая одноклеточные, колониальные и многоклеточные формы. У последних единицей структуры служит нить (трихом, или филамент). Нити бывают простые или ветвящиеся. Простые нити состоят из одного ряда клеток (однорядные трихомы), имеющих одинаковые размеры, форму и строение, или клеток, различающихся по этим параметрам.
В процессе жизненного цикла некоторые цианобактерии формируют дифференцированные единичные клетки или короткие нити, служащие для размножения (баеоциты, гормогонии), выживания в неблагоприятных условиях (споры, или акинеты) или азотфиксации в аэробных условиях (гетероцисты). Для разных представителей этой группы характерна способность к скользящему движению. Оно свойственно как нитчатым формам (трихомы и/или гормогонии), так и одноклеточным (баеоциты).
Известны разные способы размножения цианобактерии. Деление клеток происходит путем равновеликого бинарного деления, сопровождающегося образованием поперечной перегородки или перетяжки; неравновеликого бинарного деления (почкования); множественного деления Бинарное деление может происходить только в одной плоскости, что у одноклеточных форм приводит к образованию цепочки клеток, а у нитчатых — к удлинению однорядного трихома. Деление в нескольких плоскостях ведет у одноклеточных цианобактерии к формированию скоплений правильной или неправильной формы, а у нитчатых — к возникновению многорядного или однорядного трихома. Размножение нитчатых форм осуществляется также с помощью обрывков трихома, состоящих из одной или нескольких клеток, у некоторых — также гормогониями, отличающимися по ряду признаков от трихомов, и в результате прорастания акинет в благоприятных условиях.
|
|
Рассмотрим основные таксономические группы цианобактерий, различающихся морфологическими признаками (таблица 1).
Таблица 1.- Основные таксономические группы цианобактерий
| Одноклеточные формы: одиночные клетки или колонии | Многоклеточные формы: нитчатые | |||
| Пор. Chroococcales | Пор. Pleurocap- sal es | Пор. Oscillatoriales | Пор. Nostocales | Пор. Stigoneomatales |
| Размножение бинарным делением в одной или более плоскостях или почкованием | Размножение множествен- ным делени- ем или чередованием бинарного множествен- ного деления | Трихомы не- ветвящиеся, состоят из одного ряда только вегетативных клеток. Рост трихома — делением клеток в од- ной плоскости | В неветвя щихся одно рядных три- хомах помимо вегетативных образуются дифференцированные клетки: гетероцисты и иногда акинеты. Рост трихома — делением клеток в од- ной плоскости | Те же признаки, что и у представителей пор. Nostocales. Отличитель- ный признак: способ- ность веге- тативных клеток трихома к деле- нию более чем в одной плоскости приводящему к появле- нию многорядных трихомов или трихомов с истинным ветвлением , |
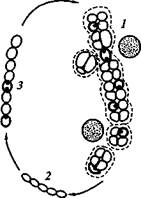
К порядку Chroococcales отнесены одноклеточные цианобактерии, существующие в виде одиночных клеток или формирующие колонии (рисунок 2). Для большинства представителей этой группы характерно образование чехлов, окружающих каждую клетку и, кроме того, удерживающих вместе группы клеток, т.е. уча-

Рисунок 2. Цианобактерии, отнесенные к порядку Chroococcales:
1 — Gloeobacter; 2 — Gloeothece', 3 — Gloeocapsa; 4 — Synechococcus; 5 — Synechocystis; 6 — Chamaesiphon. Прерывистой линией обозначены чехлы, точками — капсулы,
черточками — тилакоиды
ствующих в формировании колоний. Цианобактерии, клетки которых не образуют чехлов, легко распадаются до одиночных клеток. Размножение осуществляется бинарным делением в одной или нескольких плоскостях, а также почкованием.
В порядок Pleurocapsales выделены одноклеточные цианобактерии, способные к размножению путем множественного деления. Они существуют в виде одиночных клеток или скоплений. Скопления могут состоять всего из нескольких клеток разной формы, иметь кубическую или неправильную форму. Входящие в эту группу цианобактерии различаются способностью размножаться только множественным делением или чередованием процессов бинарного и множественного деления. Освобождающиеся в результате множественного деления баеоциты могут быть подвижными или неподвижными. Подвижность определяется отсутствием дополнительного фибриллярного слоя клеточной стенки и теряется, когда этот слой синтезируется. У неподвижных форм к моменту выхода из материнской клетки дополнительный слой клеточной стенки уже сформирован.
|
|
В последующие порядки включены многоклеточные формы цианобактерий, имеющие нитчатое строение. Особенностью цианобактерий, отнесенных в порядок Oscillatoriales, является недифференцированность трихома (последний состоит только из вегетативных клеток) и его рост путем деления клеток в одной плоскости (рисунок 3).
Для большинства представителей этой группы характерны прямые трихомы, клетки в которых дисковидные или цилиндрические, плотно прилегают друг к другу или разделены глубокой перетяжкой. Трихомы могут быть окружены общим чехлом. Скользящее движение свойственно цианобактериям, не образующим чехла или со слабым развитием последнего. К этой же группе относятся цианобактерии, имеющие подвижные спиралевидные трихомы, состоящие из клеток разной формы, не окруженные чехлом.

|
| Рисунок 3. Цианобактерии, включенные в порядок Oscillatoriales: / — Spirulina; 2 — Oscillatoria; 3 — Pseudoanabaena |

Рисунок 4. Цианобактерии, отнесенные к порядку Nostocales:
А — АпаЬаепа; Б — Nodularia; В — Cylindrospermum; Г — Nostoc, Д — Scytonema, Е — Calothrix: 1 — трихом в зрелом состоянии; 2 — гормогоний; 3 — молодой трихом. Гетероцисты изображены в виде клеток с толстой клеточной стенкой и полярными гранулами; акинеты — в виде темных клеток. Прерывистой линией вдоль трихома обозначен чехол
Дальнейший шаг по пути морфологического усложнения сделан цианобактериями, включенными в порядок Nostocales (рисунок 4). Они представлены однорядными неветвящимися нитями, рост которых происходит путем деления клеток в одной плоскости (под прямым углом к длинной оси трихома). При культивировании на среде без связанного азота некоторые вегетативные клетки дифференцируются в гетероцисты. Ряд представителей группы образует акинеты — единственный тип покоящихся форм у цианобактерий. Размножение происходит короткими обрывками трихомов, и в результате прорастания акинет, если последние образуются. У некоторых цианобактерий в дополнение к описанным выше способам размножения для этой цели служат гормогоний. Последние представляют собой короткие нити, отличающиеся рядом морфологических признаков от родительского трихома: они состоят из небольшого числа мелких активно движущихся вегетативных клеток, иногда иной формы, чем клетки родительского трихома; могут содержать газовые вакуоли; никогда не окружены чехлом. Основное отличие гормогониев от зрелых и молодых трихомов — отсутствие гетероцист.
|
|
В порядок Stigonematales объединены цианобактерии, отличающиеся от представителей предыдущего порядка способностью вегетативных клеток трихома к делению более чем в одной плоскости (рисунок 5). К этой группе отнесено несколько родов, различающихся циклами развития, расположением гетероцист в трихомах и некоторыми другими признаками.
Электронно-микроскопическое изучение вегетативных клеток цианобактерий обнаружило принципиальное сходство их строения с клетками грамотрицательных эубактерий. Более чем у 200 чистых культур определен состав оснований хромосомной ДНК. По этому признаку цианобактерий обнаруживают гетерогенность (молярное содержание ГЦ-оснований в ДНК от 35 до 71 %), сравнимую только с остальными прокариотами (25—75 %).
В качестве одной из примечательных особенностей генетического материала цианобактерий отмечают значительные различия величины цианобактериальной хромосомы. Размеры геномов, изученные более чем у 100 штаммов из разных групп, располагаются в диапазоне 1,6 — 8,6-109 Да, при этом просматривается определенная
|

Рисунок 5. Цианобактерии порядка Stigonematales:
А — Chlorogloeopsis; Б — Fischerella: 1 — зрелый трихом; 2 — гормогоний; 3 — молодой трихом. Обозначение гетероцист, акинет и чехла см. на рисунке 4
корреляция между степенью морфологической сложности и величиной генома, достигающего максимальных значений у цианобактерий со сложной организацией трихомов и циклами развития.
В группе цианобактерий сформирован самый крупный геном, обнаруженный до сих пор у прокариот. В то же время некоторые цианобактерии в отношении морфологической сложности также достигли вершины в мире прокариот и не имеют равных среди других грамотрицательных эубактерий Клетки цианобактерий, за исключением принадлежащих к роду Gloeobacter, характеризуются развитой системой внутрицитоплазматических мембран (тилакоидов), в которых локализованы компоненты фотосинтетического аппарата. Единственная энергопреобразующая мембрана Gloeobacter — цитоплазматическая, где локализованы процессы фотосинтеза и дыхания.
|
|
Большой интерес представляет группа цианобактерий из-за сосредоточения в ней разнообразных физиологических возможностей. В недрах этой группы, вероятно, сформировался и в целом оформился фотосинтез, основанный на функционировании двух фотосистем, характеризующийся использованием Н2О в качестве экзогенного донора электронов и сопровождающийся выделением О2.
У цианобактерий обнаружена способность к бескислородному фотосинтезу, связанная с отключением II фотосистемы при сохранении активности I фотосистемы.В этих условиях у них возникает потребность в иных, чем Н2О, экзогенных донорах электронов. В качестве последних цианобактерий могут использовать некоторые восстановленные соединения серы (H2S, Na2S2O3), H2, ряд органических соединений (сахара, кислоты). Так как поток электронов между двумя фотосистемами прерывается, синтез АТФ сопряжен только с циклическим электронным транспортом, связанным с I фотосистемой. Способность к бескислородному фотосинтезу обнаружена у многих цианобактерий из разных групп, но активность фиксации СО2 за счет этого процесса низка, составляя, как правило, несколько процентов от скорости ассимиляции СО2 в условиях функционирования обеих фотосистем. Только некоторые цианобактерий могут расти за счет бескислородного фотосинтеза.
Хотя подавляющее большинство цианобактерий являются облигатными фототрофами, в природе они часто находятся длительное время в условиях темноты. В темноте у цианобактерий обнаружен активный эндогенный метаболизм, энергетическим субстратом которого служит запасенный на свету гликоген, конечным акцептором электронов в которой служит О2.
Другой возможный путь получения цианобактериями в темноте энергии — гликолиз. У некоторых видов найдены все ферменты, необходимые для сбраживания глюкозы до молочной кислоты, однако образование последней, а также активности гликолитических ферментов низки. Кроме того, содержание АТФ в клетке в анаэробных условиях резко падает, так что, вероятно, жизнедеятельность цианобактерий только за счет субстратного фосфорилирования поддерживаться не может.
Цианобактерии могут ассимилировать некоторые органические кислоты, в первую очередь ацетат и пируват, но всегда только в качестве дополнительного источника углерода..
Некоторые цианобактерии способны к хемогетеротрофному росту. Набор органических веществ, поддерживающих хемогетеротрофный рост, ограничен несколькими сахарами. Это связывают с функционированием у цианобактерий в качестве основного катаболического пути окислительного пентозофосфатного цикла, исходным субстратом которого служит глюкоза.
Одна из загадок метаболизма цианобактерий — неспособность большинства из них расти в темноте с использованием органических соединений. Невозможность роста за счет субстратов, метаболизируемых в ЦТК, связана с «разорванностью» этого цикла. Но основной путь катаболизма глюкозы — окислительныйпентозофосфатный цикл — функционирует у всех изученных цианобактерий.
Цианобактерии, в группе которых, вероятно, сформировался кислородный фотосинтез, впервые столкнулись с выделением О2 внутри клетки. Помимо создания разнообразных систем защиты от токсических форм кислорода, проявляющихся в устойчивости к высоким концентрациям О2, цианобактерии адаптировались к аэробному способу существования путем использования молекулярного кислорода для получения энергии.
В то же время для ряда цианобактерий показана способность расти на свету в строго анаэробных условиях. Это относится к видам, осуществляющим фотосинтез бескислородного типа, которых в соответствии с принятой классификацией следует отнести к факультативным анаэробам.
Конструктивный метаболизм цианобактерий представляет собой шаг вперед по пути дальнейшей независимости от органических соединений внешней среды по сравнению с пурпурными и зелеными серобактериями. Цианобактерий не требуют никаких питательных компонентов в восстановленной форме. Только некоторые морские виды обнаруживают потребность в витамине В12.
Цианобактерии обладают азотфиксирующей активностью. Способность к фиксации N2, определяемая по наличию нитрогеназной активности, зависит от условий и в первую очередь от содержания в среде связанного азота и молекулярного кислорода. Основное место действия обоих факторов — нитрогеназа. В первом случае источники связанного азота репрессируют синтез и ингибируют активность фермента, во втором — О2 быстро инактивирует его.
Отрицательное действие О2 на азотфиксацию связано с восстановительной природой процесса. Так как только у цианобактерий в клетке функционируют два процесса, один из которых имеет восстановительную природу, а другой сопровождается выделением такого сильного окислителя, как О2. Возникла необходимость защиты или изолирования процесса азотфиксации от молекулярного кислорода.
Вегетативные клетки многих изученных культур обнаруживают нитрогеназную активность в анаэробных и микроаэробных условиях. Только для единичных культур, показана способность вегетативных клеток к азотфиксации в аэробных условиях, при этом до 95 % фиксированного азота приходится на темновой период, т. е. процессы фотосинтеза и азотфиксации разделены во времени. Способность подавляющего большинства цианобактерий к азотфиксации в аэробных условиях связана с гетероцистами, в которых чувствительный к О2 аппарат фиксации молекулярного азота отделен от фотосинтетического аппарата с помощью определенных ультраструктурных и биохимических перестроек..
При отсутствии в среде связанного азота некоторые вегетативные клетки (обычно 5—10%) нитчатых цианобактерий, принадлежащих к порядкам Nostocales и Stigonematales, превращаются в гетероцисты. Сформированные гетероцисты не способны к делению и не могут превращаться в вегетативные клетки.
Формирование гетероцист из вегетативных клеток сопровождается глубокими ультраструктурными и функциональными перестройками. Зрелые гетероцисты окружены тремя дополнительными слоями, внешними по отношению к клеточной стенке, что затрудняет проницаемость в них воды, ионов, нейтральных веществ гидрофильной природы и растворенных газов. Дополнительные слои, окружающие гетероцисту, в местах ее контакта с вегетативной клеткой прерываются. Перегородка, отделяющая гетероцисту от вегетативной клетки, пронизана множеством мелких каналов (микроплазмодесм), соединяющих цитоплазмы обеих клеток и обеспечивающих обмен клеточными метаболитами. В цитоплазме гетероцист в зонах контакта с вегетативными клетками располагаются светопреломляющие полярные гранулы цианофицина.
Значительную реорганизацию претерпевает в гетероцистах система фотосинтетических мембран: они укорачиваются, теряют расположение, характерное для вегетативных клеток; как правило, отмечается скопление тилакоидов вблизи полюсов гетероцисты. Морфологические изменения тилакоидов сочетаются с важными перестройками фотосинтетического аппарата на функциональном уровне. В гетероцистах не работает II фотосистема. Следовательно, внутриклеточный О2 в них не образуется. Потеря активности II фотосистемы коррелирует со следующими биохимическими особенностями гетероцист: отсутствием основных светособирающих пигментов II фотосистемы — фикобилипротеинов и содержащих их структур — фикобилисом; резко пониженным содержанием ионов марганца — необходимого компонента системы разложения воды; потерей гетероцистами способности фиксировать СО2. Деградация II фотосистемы сопровождается сохранением активности I фотосистемы, что находит отражение в поддержании значительного уровня хлорофилла а и увеличении числа реакционных центров этой фотосистемы.
В процессе формирования гетероцист наблюдается исчезновение различных цитоплазматических включений, характерных для вегетативных клеток: гликогеновых, полифосфатных гранул. В то же время в гетероцистах сохраняется в полном объеме генетическая информация, и в процессе их жизнедеятельности отмечаются активные процессы синтеза РНК и белка.
Для фиксации N2 необходим восстановитель в виде молекул восстановленного ферредоксина (иногда НАДФ • Н2) и химическая энергия в форме АТФ. Восстановитель может или непосредственно транспортироваться из соседних вегетативных клеток в готовом виде, или же генерироваться в гетероцистах в темновых ферментативных процессах из исходного транспортируемого субстрата. В последнем случае таким субстратом служит дисахарид мальтоза. Ее катаболизирование, осуществляемое по активно функционирующему в гетероцистах окислительному пентозофосфатному пути, приводит к образованию молекул НАДФ • Н2, с которых водород может передаваться на ферредоксин в реакции, катализируемой ферредоксин: НАДФ-оксидоредуктазой. Источником АТФ в гетероцистах на свету служит зависимое от I фотосистемы циклическое фотофосфорилирование, в темноте — окислительное фосфорилирование.
Нитрогеназная система катализирует восстановление N2 до аммония. Последний включается в молекулу глутаминовой кислоты в реакции, катализируемой глутаминсинтетазой:

В таком виде фиксированный азот транспортируется из гетероцист в вегетативные клетки, где с помощью глутаматсинтазы осуществляется перенос амидной группы на молекулу глутаминовой кислоты
Одна из молекул глутамата возвращается в гетероцисту для очередного акцептирования NH4, другая поступает в метаболизм вегетативной клетки.
Таким образом, все структурные и функциональные перестройки, происходящие в процессе формирования гетероцист, направлены на поддержание высокой активности нитрогеназы, что достигается, с одной стороны, путем эффективного ее снабжения восстановителем и энергией, с другой — защитой от молекулярного кислорода за счет уменьшения проникновения О2 через утолщенные оболочки гетероцист, реорганизации их фотосинтетического аппарата и высокой активности дыхания.
Прохлорофиты
Обнаружены эубактерии, осуществляющие фотосинтез кислородного типа, весьма сходные с цианобактериями, но отличающиеся от них составом фотосинтетических пигментов: отсутствием фикобилипротеинов и наличием хлорофилла b. Данные организмы названы прохлорофитами. В девятом издании определителя бактерий Берджи они выделены в порядок Prochlorales. В составе порядка 3 рода, различающихся морфологическими и некоторыми физиолого-биохимическими признаками. Это одноклеточные (сферические) или многоклеточные (нитчатые) формы, неподвижные или подвижные. Размножаются бинарным делением. Клеточная стенка грамотрицательного типа, напоминает таковую цианобактерий. Нити ДНК, не отграниченные от цитоплазмы мембраной, располагаются в центральной области клетки.
Большую часть цитоплазмы занимают тилакоиды, располагающиеся обычно концентрическими кругами по периферии клетки. Тилакоиды, как и у цианобактерий, не отделены от цитоплазмы замкнутой мембраной, и имеют тенденцию сближаться, образуя пары или стопки, состоящие из трех и более тилакоидов. Внутренние и наружные поверхности тилакоидных мембран гладкие. Фикобилисомы и фикобилипротеины не обнаружены.
Фотосинтетические пигменты представлены хлорофиллами а и b и каротиноидами. Основную массу последних составляют Р-каротин и ксантофилл. По составу жирных кислот и гликолипидов прохлорофиты близки к цианобактериям. В цитоплазме обнаружены 70 S -рибосомные частицы, содержащие РНК 16S- и 23 S -типов, аналогично рибосомальным РНК прокариот и хлоропластов эукариот. Молярное содержание ГЦ в ДНК — 39—53 %.
СО2 активно фиксируется в восстановительном пентозофосфатном цикле, о чем свидетельствуют активности двух специфических ферментов этого пути: рибулозодифосфаткарбоксилазы и фосфорибулокиназы. Конечным продуктом углеродного метаболизма на свету является полисахарид, схожий с гликогеном цианобактерий. Помимо фотоавтотрофии обнаружена способность прохлорофит к фотогетеротрофии и росту в темноте с получением энергии в процессе дыхания. Для некоторых представителей группы показана способность фиксировать N2.
Внутриклеточный О2, прохлорофиты, с одной стороны, способны использовать, обнаруживая склонность к микроаэрофилии, с другой — имеют определенные системы защиты от его токсических форм. В качестве одной из защитных систем обнаружена супероксиддисмутаза FeMn-типа, характерная для прокариотных форм.
Прохлорофиты привлекают к себе большое внимание в связи с проблемами эволюции фотосинтетического аппарата и возникновения фотосинтезирующих эукариот. Сравнение прохлорофит с цианобактериями и хлоропластами зеленых водорослей и высших растений обнаруживает черты сходства как с фотосинтетическими органеллами эукариот (организация тилакоидов, состав хлорофиллов), так и с цианобактериями (клеточное строение, состав каротиноидов, липидов, некоторые особенности метаболизма, последовательность оснований 16S рРНК). Для ответа на вопрос, в каком отношении прохлорофиты находятся с цианобактериями (развивались ли независимо и параллельно с цианобактериями, возникли ли из их предшественников, потерявших способность синтезировать фикобилипротеины, или, наоборот, цианобактерий возникли из прохлорофит), необходимо дальнейшее сравнительное изучение обеих групп эубактерии с фотосинтезом кислородного типа. Прохлорофиты рассматриваются в качестве возможных эндосимбионтов, последующая эволюция которых привела к возникновению хлоропластов зеленых водорослей и высших растений.
СПЕЦКУРС «ЧАСТНАЯ МИКРОБИОЛОГИЯ. СИСТЕМАТИКА МИКРООРГАНИЗМОВ»
|
|
|

Типы сооружений для обработки осадков: Септиками называются сооружения, в которых одновременно происходят осветление сточной жидкости...

Состав сооружений: решетки и песколовки: Решетки – это первое устройство в схеме очистных сооружений. Они представляют...

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

