Новая схема клеточной эволюции, разработанная К. Вёзе (С. Woese) на основании сравнения последовательностей нуклеотидов в рРНК малой субъединицы рибосомы, исходит из признания существующих трех фундаментально различающихся типов живых организмов: эубактерий, архебактерии и эукариот как три равноправных линии в графе отношений сходства в последовательностях рРНК. Согласно этой схеме от общего гипотетического предка, получившего название «прогенота», эволюционировали три различные ветви прокариот: эубактерий, архебактерии и уркариоты (рисунок Б). Уркариоты представлены ядерно-цитоплазматическим компонентом эукариотной клетки, включившим в себя в качестве эндосимбионтов представителей разных групп эубактерий, превратившихся в митохондрии и хлоропласты.
Эту номенклатуру предполагается заменить на узаконенную и признать существование таксона высшей категории Imperium (мн. число Imperia), а не на «надцарства». Множество живых существ распределяется между тремя империями: Eukaryota (не Eucarya); Archaeobacteria (не Archaea); Eubacteria (не Ваcteria). Соответствующие русские термины: эукариоты, архебактерии, эубактерии. Применение этой номенклатуры, помимо cоответствия правилам Кодекса, имеет то преимущество, что прокариоты имеют общий определяющий термин «bacteria», в отличии от эукариот, и таким образом закрепляется тривиальное понятие бактерий как организмов, противопоставленных композитным эукариотам. Из списка сходств и различий Eubacteria, Archaeobacteria, Eukaryota, представленных в таблице, видно, что они носят специфический характер, относящийся к деталям («коррелирующим признакам»), а не к свойствам целого организма.
Если различие между прокариотами и эукариотами очевидно, как различие между простыми (элементарными) и сложными (комбинированными) организмами, то различие между архебактериями и эубактериями далеко не столь ясно. Эубактерии и архебактерии имеют явное сходство в строении клетки, клеточном цикле, 60% сходства в последовательности рРНК; различий в метаболизме между ними не больше, чем между разными группами эубактерий (при большом сходстве в анаболизме). Сопоставление свойств ряда общих
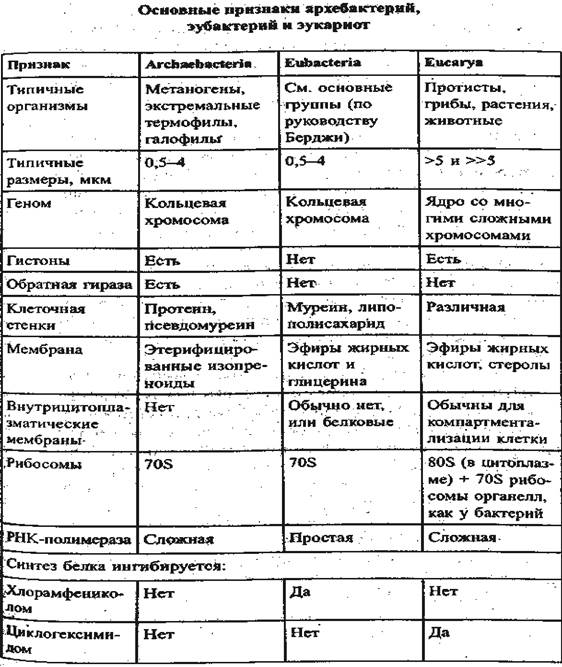
ферментов архебактерий и эубактерий, часто выполненные на отдельных представителях, без уверенности в приложимости вывода ко всему исследуемому множеству, показывает разные комбинации сходств и различий. Существенны различия в аппарате синтеза белка. Постулирование различного происхождения эубактерий и архебактерий вызывает серию вопросов о причинах сходства, на которые трудно ответить без большой серии допущений, лежащих вне филогенетического подхода. Ответ требует анализа полных последовательностей ДНК архебактерий и эубактерий и сопоставления не только генов синтеза белка, но и других.
4. Дистанционно-матричный метод построения филогенетических деревьев и их конструкции (веерообразная и сильно разветвленная дихотомическая). Гипотеза О. Кандлера о трех типах независимых проклеток
Сопоставление всех организмов по одному гену малой рибосомальной РНК, 16S для прокариот и 18S для эукариот, дало возможность построить единую классификацию всего живого мира, а не переходить к разным наборам признаков, как это неизбежно при сравнении, например, насекомых и грибов. Анализ по 16S ДНК предоставил возможность создать единую классификационную систему, которую можно отразить в виде графа сходства последовательностей оснований в гене. Последовательность оснований устанавливается аналитически с большой точностью. Сопоставление последовательностей отчасти зависит от взятого для анализа набора организмов, и здесь могут получаться разные деревья.
Следующим вопросом было сопоставление этого классификационного дерева с эволюцией. Постулировалось, что мутации, ведущие к замене нуклеотидов, происходят случайно и число замен находится в прямой зависимости от времени; наибольшее число замен должно накопиться у самых древних организмов. Приняв это допущение, можно реконструировать на основе классификационного дерева рРНК-гена филогенетическое дерево организмов. Результат такой реконструкции в общем совпал с картиной эволюции, установленной на основе палеонтологии и сравнительного подхода: многоклеточные растения, животные, грибы заняли вершину дерева. Упорядочение множества прокариот было субъективным, и те совпадения в группировании, которые обнаружились, например, у грамотрицательных и грамположительных бактерий, послужили основанием для принятия новой систематики как естественной. Следующим вопросом является соотношение филогении и эволюции. Оба понятия подразумевают последовательность во времени, но филогения относится к генеалогии организма или групп организмов, а эволюция может подразумевать и более широкие понятия, например эволюцию осадочного процесса, который сохраняет свою историю в виде последовательных слоев. Твердо установленные палеонтологические данные имеют силу факта, а филогения - реконструкции. Все эти соображения становятся особенно важны, когда рассматриваются наиболее простые организмы — прокариоты. Молчаливо допускается, что древние организмы сохраняются без существенных изменений.
Различия во внутренней сфере организма могут иметь слабое выражение в его направленных к среде обитания функциональных или фенотипических характеристиках. Для экосистемы неважно, какие внутренние изменения происходят в клетках и аппарате синтеза белка, коль скоро он функционирует, но очень важны химические функции организма. При анализе противопоставляются два подхода: филогенетических последовательных рядов и комбинаторный для сопоставления фенотипических функций.
Установление последовательностей оснований в рРНК привело к возможности немыслимой ранее идентификации прокариотных организмов и положения их на филогенетическом дереве без определения фенотипических свойств. Иерархическая система устанавливается благодаря соглашению относительно условных критериев рангов таксонов по числу замен оснований
Эубактерии распределились по следующим главным ветвям
16S рДНК филогенетического дерева, представленном на рисунке 2
как дихотомический ключ.
Труднее всего сопоставить филогенетическую и фенотипическую систематики. В одни и те же филогенетические ветви попадают крайне разнообразные по физиологии организмы, например, анаэробные фотоавтотрофы пурпурные бактерии и эубактерии, цитофаги и бактероиды. При переходе от кpyпных ветвей грамположительных и грамотрицательных бактерий к низшим иерархическим уровням таких несовпадений становится все больше: Из приведенных на схеме групп бактерий подавляющее большинство таксонов входит в группировки цианобактерий, протеобактерий, грамположительных

Рисунок 2. Основные ветви филогенетического дерева эубактерий
организмов. Эти три ветви хорошо различимы. Самостоятельное положение ветвей спирохет, зеленых фототрофов также согласуется с традиционным представлением о единстве и своеобразии этих групп. Зеленые бактерии обычно функционируют в сходной эконише с пурпурными серными, но сильно различаются по строению фотосинтетического аппарата. Однако объединение цитофаг, традиционно считавшихся близкими к аэробным миксобактериям, и облигатно анаэробных бактериоидов не имеет функционального обоснования.
Наибольшую трудность в корреляции филогенетической и
функциональной систематики представляет наиболее многочисленный и разнообразный комплекс цианобактерий, протеобактерий, грамположительных организмов. Было сделано предположение Г. А. Заварзиным и др., что этот комплекс отражает свое происхождение из циано-бактериального сообщества как стволовой эволюционной группировки микроорганизмов. Для грамотрицательных организмов вместо обозначения «пурпурные бактерии и их родственники» был предложен класс Prоteobacteria. Протеобактерий, ветви которых оцениваются в ранге подклассов, пока обозначаемых греческими буквами, попытались разделить на экологические группировки. Альфа-подкласс включает группу пурпурных несерных бактерий, почкующиеся и простекобактерии, ризобии и агробактерии; среди органотрофов этого подкласса относительно много олиготрофов и организмов, связанных с высшими растениями. В бета-подкласс входят «газотрофы», например, метанотрофы и нитрификаторы, составляющие естественную группу по особенностям строения клетки с мощным мембранным аппаратом и обменом, основанным на окислении газов анаэробного происхождения. Гамма-подкласс включает группы органотрофных копиотрофов: факультативных анаэробов-бродильщиков (энтеробактерий и вибрионов), типичных аэробных окислителей (псевдомонад), а также бесцветные серобактерии, серные пурпурные бактерии: Корреляция филогенетических групп протеобактерий с экологическим положением в системе носит характер лишь общей тенденции с многочисленными отклонениями.
Последующее дробление свойственно всем способам классификации и ведет к превращению идентификационных деревьев в «кусты». Примером может служить иммунодиагностика энтеробактерий, которая привела в свое время к избыточному числу субродовых таксонов сальмонелл. Это общая судьбам классификационных схем, которую они претерпевают при дальнейшей детализации.
Неудобством классификации по 16S РНК оказывается появление случаев, когда фенотипически сходные организмы оказываются принадлежащими разным ветвям, и возникает дробление родов по единственному критерию с возрастающей неустойчивостью систематики на родовом уровне. Из положения организма на филогенетическом дереве можно сделать лишь ограниченные заключения о его функции. В качестве общего представления можно полагать, что прокариоты имеют некоторые сердцевинные компоненты, такие как репликация генома и синтез белка, и периферические, определяющие их функции, значимые для занятия функциональной (экологической) ниши выживания в течение эволюции.
Грамположительные организмы, таксономия которых находится в стадии формирования, разделились на ветви с «низким Г + Ц» и «высоким Г + Ц».
Коротко следует перечислить малые ветви филогенетической
системы. Ближе всего к корню дерева собственно бактерий полагаются экстремально термофильные водородные бактерии родов Hydrpgenobacter (Calderobacterium), Aquifex — хемолитоавтотрофы с необычным путем ассимиляции углекислоты, paстущие при температуре выше 70 °С за счет окисления Н2 и очень богатые цитохромом.
Следующую группу составляют экстремально термофилиные органотрофные Thermatoga, грамотрицательные организмы, имеющие на концах клеток характерные вздутия наружной мембраны, создающие пузыри перипласта. Термотоги относительно легко выделяются из разных мест обитания, как морских, так и пресноводных, а также из подземной гидросферы
Нитчатые фотогетеротрофные зеленые хлорофлексусы обычно обнаруживаются в термальных источниках при более высокой температуре, чем термофильные цианобактерии.
Дейнококки в общем напоминают аэробные органотрофные микрококки, но отличаются необычайной радиоустойчивость.
Цитофаги и бактероиды составляют многочисленные и важные группы и обсуждаются no-отдельности, не имея общих фенотипических свойств. Цитофаги служат важнейшими аэробными деструкторами целлюлозы и, благодаря их способности к скольжению, долго рассматривались как не имеющие плодовых тенл миксобактерии. Бактероиды — пептолитические анаэробы, составляющие основную часть бактериального населения кишечника.
О. Кандлер, К. Вёзе, и К. Штетгер нашли, что наиболее уклоняющиеся по последовательностям рРНК («древние») группы принадлежат экстремальным термофилам и на этом основании предположили происхождение, прокариот из гидротермальных мест обитания.
Специализация архебактерий позволяет рассматривать отдельно их функциональные группы — метаногенов, галофилов, экстремальных термофилов. Роды экстремальных термофилов, вследствие своего уникального положения на 16S рРНК дереве, получают высокие таксономические ранги.
Заканчивая обзор филогенетических ветвей прокариот, следует заметить, что он дает очень мало опорных пунктов для логической систематизации сведений, а тем более руководящих принципов в построении большой системы, филогенетическая система, основанная на одном рибосомальном гене, не более чем одна из технически удобных и разработанных систем упорядочения множества организмов с целью их идентификации.
Итак, в настоящее время отсутствует сколько-нибудь детализированная эволюционная система прокариот. Все описанные выше попытки подойти к ее созданию позволяют сделать вывод о том, что решение этой проблемы - дело неблизкого будущего. Особенности прокариот в области морфологической, физиолого-биохимической, генетической организации говорят о неприменимости к ним хорошо разработанных принципов, используемых при построении системы высших организмов. Это, естественно, значительно усложняет задачу, но не делает ее безнадежной. Уже сейчас в мире прокариот для наиболее изученной части этих организмов - эубактерий можно проследить основные направления эволюционного развития. Одна из многообещающих идей заключается в том, что в основе прогрессивной эволюции эубактерий лежит совершенствование способов получения ими энергии.








