

Историки об Елизавете Петровне: Елизавета попала между двумя встречными культурными течениями, воспитывалась среди новых европейских веяний и преданий...

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Историки об Елизавете Петровне: Елизавета попала между двумя встречными культурными течениями, воспитывалась среди новых европейских веяний и преданий...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Топ:
Выпускная квалификационная работа: Основная часть ВКР, как правило, состоит из двух-трех глав, каждая из которых, в свою очередь...
Основы обеспечения единства измерений: Обеспечение единства измерений - деятельность метрологических служб, направленная на достижение...
Процедура выполнения команд. Рабочий цикл процессора: Функционирование процессора в основном состоит из повторяющихся рабочих циклов, каждый из которых соответствует...
Интересное:
Финансовый рынок и его значение в управлении денежными потоками на современном этапе: любому предприятию для расширения производства и увеличения прибыли нужны...
Искусственное повышение поверхности территории: Варианты искусственного повышения поверхности территории необходимо выбирать на основе анализа следующих характеристик защищаемой территории...
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
ЯПК состоят из нуклеопоринов.

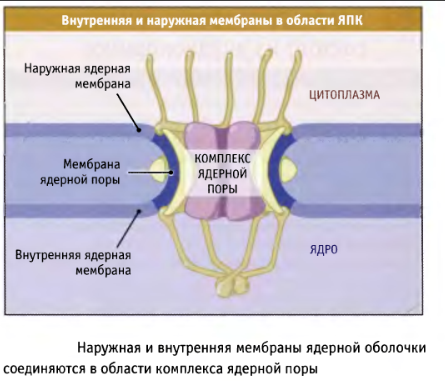
Транспортёр (втулка) - часть ЯПК, наиболее прочно связывающаяся с карго, транспортируемым через канал. Либо комплекс карго с рецептором.
Карго - любая макромолекула, транспортирующаяся из одного компартмента клетки в другой.
Окончатые мембраны - стопки двойных мембран в цитоплазме, содержащих ЯПК.


(сайты причаливания для транспортных рецепторов)

Ещё одна особенность нуклеопоринов - присутствие суперспирали (биспирали) - α-спирали, связывающейся с другими α-спиральными доменами с образованием мультимерной структуры.

Белок, не содержащий адресного сигнала, может транспортироваться, присоединившис к белку, имеющему сигнал.
Обычно NLS (nuclear localization signal) содержит лизин и аргинин.
Двойной сигнал: NLS содержит две коротких последовательности основных а/к, разделённых примерно 12 различными а/к. [у нуклеоплазмина - негистонового внутриядерного белка]





Вместе с другими транспортными ядерными факторами импортин β играет роль в сборке митотического веретена и ЯПК, а также в формировании ядра после завершения митоза.

Ядерно-цитоплазматическая циркуляция белков. Церкулирующие белки содержат NLS и NES ( nuclear export signal - сигнал ядерного экспорта).

Экспортин - рецептор ядерного экспорта, представитель семейства кариоферинов. Основной экспортин представлен белком Crml .


Белок Rev содержит информацию об импорте в форме классического NLS , и отдельно информацию об экспорте ( NES ).
Мутанты Rev не поддерживали репликацию вирусов.


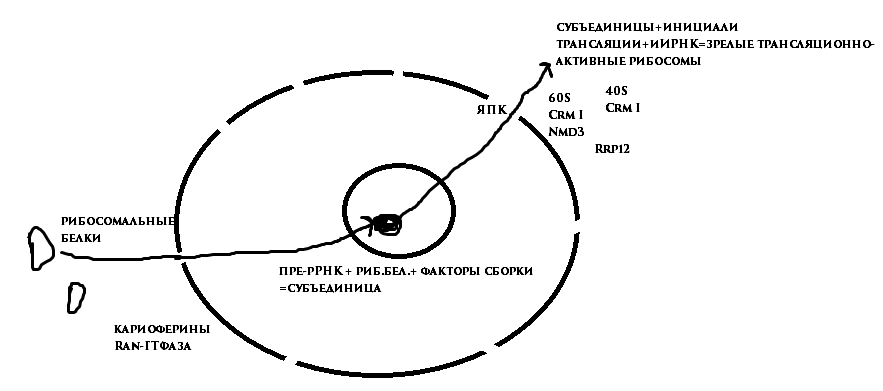
Рибосомы - большие нуклеопротеидные комплексы из двух субъединиц, содержащих около 80 белков и 4 молекулы рРНК. Они начинают собираться по отдельности в ядрышке и для окончательной сборки экспортироваться в цитоплазму. Субъединицы рибосом - одни из самых крупных комплексов, транспортируемых через ЯПК.
|
|
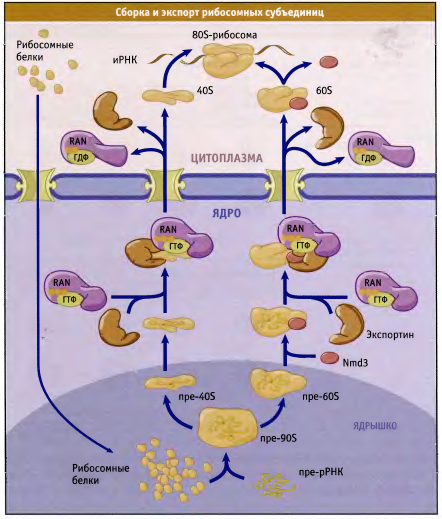
В ядрышке образуются пре-рРНК. В цитоплазме образуются рибосомальные белки, импортируются в ядро благодаря кариоферинам и Ran -ГТФазе, попадает в ядрышко, связывается с пре-рРНК и факторами сборки. Образованные субъединицы экспортируются из ядра через ЯПК. Попав в цитоплазму, связываются с инициалями трансляции и иРНК, образуя зрелые трансляционно-активные рибосомы. Экспорт субъединиц рибосом: с большой субъединицей (60 S ) связывается адаптерный белок Nmd 3, способствующий присоединению Crm I; с малой субъединицей Nmd 3 НЕ связывается, адаптер пока не обнаружен, присоединяется Crm I. Другая группа исследований, тоже проведённых на дрожжах, показывает участие в экспорте белка Rrp 12, который структурно близок к семейству кариоферинов и, как и все они, связывается с FG -повторами нуклеопоринов и с Ran , а также с пре-рРНК.

тРНК образуются в ядре, где они подвергаются процессингу и затем выходят в цитоплазму.

НАПИСАТЬ ТЕОРИЮ!!!!!!!!!!!!!!
С. 272

ДОЧИТАТЬ ПРО ИРНК!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!
Метод FISH.
С помощью флуоресцентной микроскопии также можно локализовать в клетках молекулы нуклеиновых кислот или их фрагменты. Для этого используется метод FISH (метод гибридизации флуоресцирующих молекул нуклеиновых кислот). В этом случае на клетки с денатурированной ДНК в составе ядер или хромосом наносят меченные флуорохромами однонитчатые молекулы нуклеиновых кислот, которые в процессе ренатурации связываются только с комплементарными последовательностями ДНК на препарате и таким образом четко выделяют их среди других молекул ДНК в ядрах.
Используют короткие последовательности ДНК (зонды), которые являются комплементарными по отношению к исследуемым последовательностям ДНК. Зонды гибридизуются (связываются) с комплементарными участками ДНК и благодаря тому, что они помечены флуоресцентной меткой, позволяют видеть локализацию интересующих генов в составе ДНК или хромосом. В отличие от других методов изучения хромосом, требующих активного деления клетки, FISH можно выполнять на неделящихся клетках, благодаря чему достигается гибкость метода.
|
|
FISH может применяться для различных целей с использованием зондов трех различных типов:
39. Понятие кариотипа.
Кариотип — хромосомный набор, совокупность признаков хромосом.
Кариоти́п — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы).
Внешний вид хромосом существенно меняется в течение клеточного цикла: в течение интерфазы хромосомы локализованы в ядре, как правило, деспирализованы и труднодоступны для наблюдения, поэтому для определения кариотипа используются клетки в одной из стадий их деления — метафазе митоза.
Для процедуры определения кариотипа могут быть использованы любые популяции делящихся клеток. Для определения человеческого кариотипа используют, как правило, лимфоциты периферической крови, переход которых от стадии покоя G0 к пролиферации провоцируют добавлением митогена фитогемагглютинина. Для определения кариотипа могут быть использованы также клетки костного мозга или первичная культурафибробластов кожи. Для увеличения числа клеток на стадии метафазы к культуре клеток незадолго перед фиксацией добавляют колхицин или нокадазол, которые блокируют образование микротрубочек, тем самым препятствуя расхождению хроматид к полюсам деления клетки и завершению митоза.
|
|
После фиксации препараты метафазных хромосом окрашивают и фотографируют; из микрофотографий формируют так называемый систематизированный кариотип — нумерованный набор пар гомологичных хромосом, изображения хромосом при этом ориентируются вертикально короткими плечами вверх, их нумерация производится в порядке убывания размеров, пара половых хромосом помещается в конец набора.
Для изучения ультраструктуры хромосом широко применяется метод получения целых выделенных митотических хромосом. На таких препаратах видно, что в состав хромосом входят элементарные фибриллы диаметром 25-30 нм. Однако, уловить характер укладки, какой-либо порядок в их расположении не удается. Хромосомы в этом случае имеют вид тел, состоящих из перепутанных изгибающихся фибрилл.
Хромосомы включают три компонента – собственно тело (плечо), теломерный участок и центромеру. Это палочковидные структуры, разной длины, обычно имеют два плеча, соединенных в зоне центромеры – первичная перетяжка. Плечи заканчиваются теломерами. Если плечи равны – мета, не равны – субмена, одно плечо своем не заметно – акро, сразу за центромерой теломера – тело. В области первичной перетяжки есть кинетохор. К нему присоединяются микротрубочки во время митоза. Хромосомы могут содержать несколько центромер. Там же присутствует особая сателлитная ДНК. У некоторых хромосом есть вторичная перетяжка – ядрышковый организатор. Она расположена около дистального конца, отделяет небольшой участок (спутник). Теломерные концы не могут соединяться с другими хромосомами, там есть особая ДНК – защищающая хромосому от укорачивания в процессе синтеза.
|
|
Митотические хромосомы – конденсированы, не теряют целостности даже при удалении гистонов. Состоят из рыхлой сети фибрилл и белковых петель. Хроматин особым способом компактизуется (уровни упаковки). Они приводят к структуре хромосомы.
 ????????????
????????????
ПОЛИТЕННЫЕ ХРОМОСОМЫ
· Интерфазные хромосомы, максимально декомпактизованы, гены имеют максимальные возможности для экспрессии - активно функционирующие хромосомы;
· Имеют гигантские размеры, т.к. состоят из тысяч гомологичных нитей - хроматид;
· Характерная поперечная исчерченность - рисунок хромомеров;
· Число хромосомных элементов в ядрах с политенными хромосомами чаще всего гаплоидное, т.к. гомологичные хромосомы одной пары тесно конъюгируют друг с другом, в рез-те чего общее число хромосом уменьшается вдвое.
Явление соматического синапсиса состоит в том, что гомологичные хромосомы каждой пары объединяются. Оба элемента конъюгируют диск к диску максимально точно, так что создается впечатление единой хромосомы. В результате этого число политенных хромосом в ядре уменьшается до гаплоидного.
Синапсис может быть нарушен. При асинопсисе хромосомы не конъюгируют, а располагаются в ядре раздельно. Частота асинапсиса может значительно изменяться под действием различных факторов, к примеру, варьирование температуры.
Поперечная исчерченность. Вдоль каждой хроматиды располагаются участки более (хромомеры) и менее (межхромомеры) плотной упаковки ДНП. Когда многочисленные сестринские хроматиды тесно конъюгируют, гомологичные хромомеры сближаются и образуют поперечную полосу (диск). Деконденсированные участки хроматид между хромомерами в политенной хромосоме образуют междиски. Рисунок дисков уникален.
Клеточный цикл политенных хромосом состоит из двух фаз: S (синтетическая), G (межсинтетическая).
Выделяют два типа расположения хромосом в ядре: независимое друг от друга и связанное: с объединением прицентромерных районов всех хромосом в общий хромоцентр.
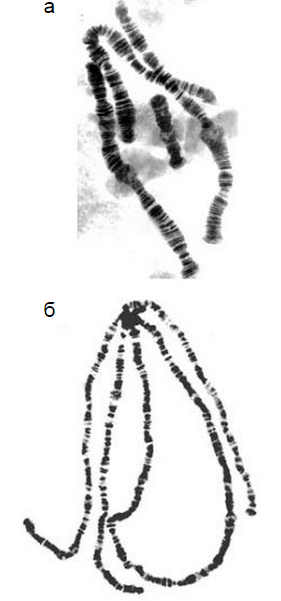
Если коньюгация гомологичных хроматид максимальна, образуются “классические” политенные хромосомы, т.е. цилиндрические жгуты с отчетливым рисунком дисков (А). Если степень коньюгации хроматид минимальна, образуется полиплоидное ядро с ретикулярной структурой. Это так называемая скрытая политения (Б). В некоторых случаях коньюгация хроматид нарушается очень сильно, но только в некоторых питающих клетках (С).
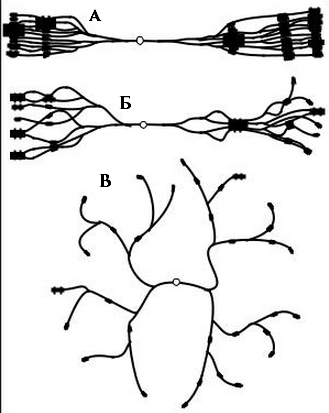
К настоящему времени имеются многочисленные и разнообразные доказательства того, что хромосомы этого типа являются пучком индивидуальных нитей - хроматид. Самым простым доказательством политенности хромосом является, повидимому, измерение количества ДНК в ядрах с политенными хромосомами и отнесение этого количества к содержанию ДНК в обычном диплоидном или гаплоидном ядре. Это отношение будет характеризовать степень плоидности (политенности) хромосом в данном типе клеток на данной стадии развития. Для этого чаще всего используют методики цитофотометрии или спектрофотометрии. Использование этих методов основано на измерении поглощения света определенной длины волны при прохождении через окрашенный или неокрашенный препарат.
|
|
Политения возникает и достигает высоких степеней в тканях, органах или на тех стадиях развития, когда есть необходимость быстрого развития органа при неизменно высоком уровне функционирования. Органы, содержащие клетки с политенными хромосомами, как правило вовлечены в процессы интенсивной секреции, осуществляемые в течение короткого времени на фоне быстрого роста. Таковы питающие клетки ооцитов, глоточные, шелкоотделительные, слюнные железы, трофобласт, антиподы, гаусторий, суспензор, эндосперм и т.д.
Гигантские вздутия - пуфы - формируются из дисков, а затем вновь превращаются в диски.
http://www.studfiles.ru/preview/5136163/page:2/
И Т.Д.!!!!!!!!!!!!!!!!!!


Липидозы - наследственные заболевания НС, обусловленные растройством обмена липидов.
Липодистрофия - патологическое состояние, характеризующееся зачастую патологией жировой ткани. Встречается два основных вида липодистрофии: атрофия и гипертрофия.
Атрофия - растройство питания органов. Гипертрофия - увеличение объёма и масса органа.
Дислипопротеинемии — количественные и качественные нарушения состава липопротеинов крови. Основными Д. являются гипер- и гиполипопротеинемии
ЦЕНТРИОЛИ 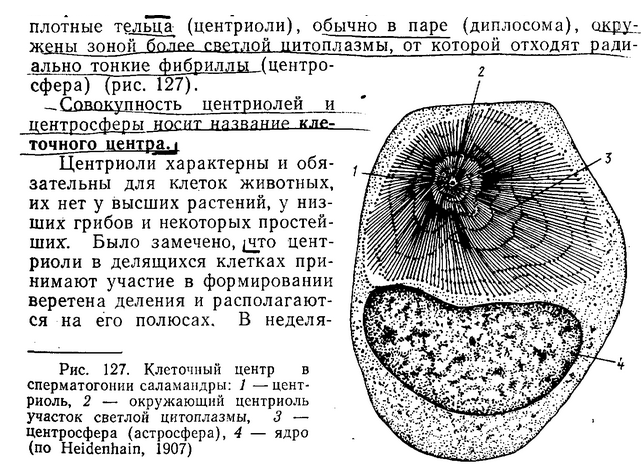


Центриолярный цикл.
Центриолярный – дупликация цетриолей. Центросомный = центриолярный + уменьшение перецентриомерного материла.
В митозе цетросомы имеют сходное строение, несмотря на то, что за время митоза происходит ряд существенных перестроек: конденсация хромосом, разрушение ядерной оболочки, образование веретена деления, расхождение хромосом. В митозе в клеточных центрах (их два, по одному на каждом полюсе) находится по диплосоме. Дочерняя центриоль своим концом направлена на материнскую. Материнская центриоль на всех стадиях митоза окружена довольно широкой зоной (0,3 мкм) тонких фибрилл – центриолярное фибриллярное гало. От этого гало радиально отходят микротрубочки. У дочерних центриолей нет ни гало, ни МТ. В это время происходит формирование веретена митотического аппарата, состоящего из МТ. На концах веретена располагаются диплосомы, окруженные радиальными микротрубочками (центросфера). В данном случае можно говорить о том, что клеточные центры являются центрами организации микротрубочек. Доказательство – колхицин, т.е. если воздействовать на клетку холодом или колхицином, мт исчезнут, а затем возникнут главным образом в районе материнских центриолей диплосом, в каждом из полюсов клетки. Мт начинают отрастать от зоны гало, расположенной на материнской центриоли. Важно отметить, что в это время на материнских центриолях нет сателлитов, и в это же время цитоплазма теряет микротрубочки: МТ цитоплазмы разбираются, а пул освободившихся тубулиновых мономеров идет на образование МТ веретена и центросферы, которые образуются на фибриллярном гало, как на ЦОМТ. К телофазе толщина фибриллярноо гало уменьшается.
К концу телофазы, когда произошло разделение клетки надвое, а хромосомы начали деконденсироваться и образовывать новые интерфазные ядра, идет разрушение веретена деления, его МТ деполимеризуются. Клеточные центры меняют свою структуру. Материнская и дочерняя центриоли теряют взаимно перпендикулярное расположение, отходят на небольшое расстояние. Вокруг материнской цетриоли гало и МТ не выявляются. В начале G1-периода на поверхности материнской центриоли возникают сателлиты, имеющие ножку и головку, от которой радиально отходят МТ, начинают расти в длину и заполнять собой цитоплазму. Следовательно, вторая форма активности клеточного центра – образование цитоплазматических МТ в интерфазных клетках. Если считать клеточные центры основными местами образования МТ, то общее количество последних должно быть равно числу МТ, отходящих от центриолей. От клеточных центров в интерфазе отходит всего лишь несколько десятков МТ, а в цитоплазме их так много, что с помощью иммунофлуоресцентного метода их трудно подсчитать. По мере роста МТ, часть из них теряет связь с областью центриолей и может находиться в цитоплазме долгое время.
Если клеткам запретить переходить в S-период, они могут существовать в фазе клеточного покоя (G0-период). В это время материнская центриоль продолжает функционировать как центр образования МТ цитоскелета. Но одновременно она может проявлять еще одну форму активности – образовать ресничку (аксонема образуется из A и B -мт). Это – третья форма активности центриолей как ЦОМТ. При наступлении S-периода клеточный центр приступает к четвертой форме активности: происходит удвоение числа центриолей. В это время около каждой из разошедшихся еще в конце телофазы центриолей, идет закладка новых центриолярных цилиндров – процентриолей. В районе проксимальных концов каждой центриоли перпендикулярно длинной оси закладывается 9 одиночных МТ, затем они преобразуются в дуплеты, а потом в триплеты, растущих от новых цилиндров.
S -период. Около каждой (и мат., и доч.) центриоли закладываются новые центриолярные цилиндры - процентриоли. (9 синглетов (одиночных)--> 9 дуплетов --> 9 триплетов мт). Закладка процентриолей происходит на проксимальных концах. В этом месте растут новые поколения центриолей, тоже с проксимального конца. Во время роста процентриолей здесь можно видеть центральную «втулку» со спицами.
Благодаря такому росту структур образуется сначала короткая дочерняя центриоль, т.е. процентриоль, которая затем дорастает до размера материнской, . этот способ увеличения центриолей – редупликация. Важно отметить, что размножение центриолей не связано с делением, почкованием или фрагментацией, а идет путем образования зачатка процентриоли.
Центросомный цикл.
Центросомный = центриолярный + уменьшение перицентриомерного материла.
Клеточный центр (центросома) состоит из двух центриолей и периценриолярного материала.
В S-периоде во время удвоения центриолей, материнская центриоль проявляет вторую форму активности, она продолжает быть центром образования МТ. В результате процесса дупликации около каждой центриоли вырастает новая дочерняя центриоль. Поэтому в клетке после завершения S-периода находятся две диплосомы.
После этого наступает следующий период клеточного цикла – постсинтетический G2 период, когда в клетке начинается подготовка к делению. Исчезают сателлиты на материнской диплосоме, а обе материнские центриоли покрываются фибриллярным гало. Происходит исчезновение МТ.
Нарушения центриолярного цикла могут вызвать ряд патологических изменений клеток, в первую очередь появление многополюсных митозов. Так, при действии бетта-меркаптоэтанола происходит блокада нормального митоза, при этом диплосомы расходятся на отдельные центриоли. При отмывании от этого вещества клетка снова приступает к делению, но в этом случае каждая центриоль активируется и образует полюс веретена. Таким образом, возникают трех- и четырехполюсные митозы, обусловливающие неравномерное распределение хромосом. Это приводит к анеуплоидии, которая вызывает гибель клетки.
В подавляющем большинстве клеток, центросомы участвуют в полимеризации тубулинов и являются структурами, играющими роль центров организации МТ. Микротрубочки самих центриолей служат затравками для полимеризации тубулинов только в одном случае – при росте аксонемы реснички. Это временное состояние. При переходе клеток к делению, реснички могут исчезать, а базальное тельце снова выполнять функции центриоли, участвуя в организации цитоплазматических МТ и Мт веретена. Только в этих случаях, центрами оранизации МТ являются не сами центриолярные цилиндры, а перицентриолярный материал (головка сателлитов, олколоцентриолярный матрикс, гало и т.д.) Следовательно, центриоль нужно рассматривать как один из компонентов более сложной структуры – центросомы. Это оговорка связана с тем, что у всех растений ЦОМТ не содержит центриолей. Более того, в раннем эмбриогенезе позвоночных животных образуется веретена деления, не имеющие центриолей в полюсах.
Процесс образования веретена деления: мт, растущие от центриолей, как бы заталкивают хромосомы в экваториальную плоскость. Полное оформление веретена создаётся за счёт взаимодействия как мт центриолярного (или просто полюсного происхождения в случае клеток ВР), так и кинетохорного происхождения.

Белки центриолей: ɤ-тубулин, центрин, перицентрин, p 210 и др.
Хромосомы тоже участвуют в образовании веретена. Белок RAN - GTF - мощный центр нуклеации мт. В основном локализован вблизи хромосом, т.к. там есть RAN - GEF (функция: RAN - GDF --> RAN - GTF ). Мт растут от тела и плеч хромосом.Эти МТ захватываются кинетохором. т.к. их концы после нуклеации отсоедин от хромосомы, то кинетохор захватывает + концы. «–» конец такой МТ с динеином торчит в цитоплазму. При его вз-ии с «+» концом полюсной МТ динеин в качестве карго повезет МТ. (т.о. хромосома будет двигаться к полюсу). Значит, веретено деления м.б. образовано вовсе без участия центриоли.
ЦЕНТРОСОМА

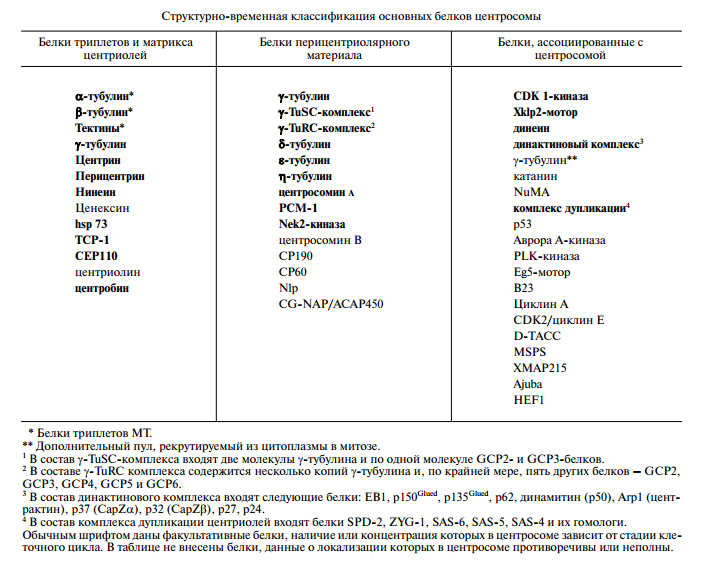
Белки триплетов центриолей.
Тектины ( tektins ) - белки, постоянно присутствующие в центриолях, ресничках, жгутиках, БТЖ. Тектин-А, тектин-В, тектин-С. В составе жгутика тектины формируют тонкие филаменты.
Белки матрикса центриолей.
Центрин, или кальтрактин ( centrin , caltractin ) - кислый белок матрикса.Формирует филаменты. Частично содержится в центросоме, а большая часть - в цитоплазме.
Перицентрин ( pericentrin ) - интегральный белок центросомы. Перицентрин-А, перицентрин-В (кендрин).
Нинеин ( ninein ) - самозакрученный кислый белок. Имеет сайт связывания GTP. Преимущественно обнаруживается на материнской центриоли.
Ценексин ( cenexin ) - на одной из центриолей.
|
|
|

Архитектура электронного правительства: Единая архитектура – это методологический подход при создании системы управления государства, который строится...

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...

Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!