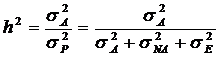Как уже отмечалось ранее, изменчивость в популяциях может быть разделена на генотипическую и средовую компоненты. Простая модель для оценки фенотипа индивидуального дерева может быть выражена (11.2) как:
P = G + E,
или фенотип = генотип + влияние среды.
Эта простая модель для индивидуальных оценок дерева может быть расширена до оценки изменчивости популяции индивидуумов как (11.1), или:
s 2 = s 2 G + s 2 Е,
то есть, фенотипическая изменчивость = генотипическая изменчивость + средовая изменчивость.
Генотипическая изменчивость в таком случае может быть разделена на аддитивные и неаддитивные компоненты:
s G 2 = s 2 А + s 2 NA (14.6).
А модeль общей фенотипической изменчивости может быть представленa как:
s Р 2 = s 2 А + s 2 NA + s 2 Е (14.7)
Таковы общие теоретические представления, но как получить эти оценки в действительности? Лучший путь оценить генетическое преимущество какого либо дерева – это сравнить его потомство с потомствами других деревьев. Такие генетические опыты предпринимаются, чтобы отделить генотипические от экологических различий, выращивая все потомства в сходных условиях местопроизрастания. Следовательно, если родитель А имеет более высокое потомство, чем родитель В в сходных условиях местопроизрастания, и эти различия могут быть подтверждены статистически, то говорят, что родитель А производит генетически улучшенное потомство (по определенному признаку). Главные усилия в большинстве селекционных программ предпринимаются для установления и ранжирования отобранных фенотипов по их генетическим качествам или ценности.
Генетическая ценность (оценка) родительских деревьев выражается в терминах комбинационной способности. Существует два типа комбинационной способности, представляющих интерес для лесных селекционеров: общая комбинационная способность и специфическая комбинационная способность.
Общая комбинационная способность (ОКС или G CA – general combining ability) представляет среднюю оценку отклонения потомков данного генотипа от средних всех полученных гибридов всех генотипов, включенных в опыт. По определению Д. С. Фолконера (1985), общая комбинационная способность «представляет собой среднее значение по всем F 1, для которых данная линия является родительской, и ее величина выражается как отклонение от общего среднего по всем кроссам». Тогда любой конкретный гибрид или кросс имеет ожидаемое значение, равное сумме общих комбинационных способностей родительских линий. Результат, однако, может отличаться от ожидаемого значения в той или иной мере. Это отклонение называется специфической комбинационной способностью (СКС или SCA – specific combining ability).
В статистических терминах общие комбинационные способности соответствуют главным эффектам, а специфические - взаимодействиям.
Для уяснения существа комбинационных способностей рассмотрим пример, приводимый в книге B. Zobеl & J. Talbert (1984).
Указанные авторы анализируют результаты скрещивания между восемью отобранными деревьями с целью установления их генетической ценности. Четыре из них выбраны в качестве отцовских родителей и четыре – в качестве материнских. Каждое мужское скрещивалось с каждым женским, а потомство высажено в испытательные культуры. Через несколько лет после полевых испытаний потомство было оценено. Средние величины каждого кросса были измерены в единицах объема и представлены в виде таблицы (табл. 14.2). В ней указаны так же средние величины потомства каждого родителя и средняя величина потомства опыта в целом.
Можно заметить, что средняя величина потомства отдельного взятого скрещивания (кросса) деревьев (5 х 1) равна 9 объемным единицам, в то время, как средняя величина потомства от всех скрещиваний родительского дерева № 5 равна 13-ти объемным единицам. Средняя величина по опыту для всех потомств деревьев равна также 13-ти объемным единицам.
Общая комбинационная способность (ОКС) определяется как средняя величина потомства отдельного дерева, когда оно скрещивается со множеством других деревьев.
Хотя показатели ОКС могут быть выражены в абсолютных единицах, обычно считается более удобным и осмысленным выразить их как отклонения от общей средней. То есть, ОКС дерева – это способность его давать при скрещивании с другими деревьями определенный средний уровень развития признака. ОКС будет выше у того дерева, у которого среднее значение признака потомства выше, чем у других. Таким образом, родитель с ОКС, равной 0, имеет среднюю ОКС. Положительная ОКС указывает на родителя, который производит потомство выше среднего, в то время как отрицательная ОКС - на родителя, производящего потомство, которое является ниже среднего для всей популяции.
Данные, представленные в таблице (14.2), использовались для расчета ОКС каждого из родителей. Например, ОКС отцовского дерева № 2 рассчитано как:
ОКС2 = средняя величина дерева № 2 – средняя по опыту = 17-13 = + 4.
Дерево № 2, следовательно, имеет ОКС2 по объему + 4 единицы. Другие ОКС могут быть рассчитаны таким же образом. Например, ОКС для дерева № 4 равна + 1 объемной единицы, в то время, как ОКС дерева № 3 – 3 единицы.
Селекционная ценность каждого дерева определяется как удвоенная ОКС. Различие между селекционной ценностью и ОКС понятно из природы этих показателей. Селекционные ценности являются бóльшими, ввиду того, что рассматриваемый родитель вкладывает только половину генов в свое потомство, другая половина приходит от других членов популяции. Так, селекционная ценность родителя № 2 может быть рассчитана как:
2 (ОКС2) = 2 (4) = 8.
Таблица 14.2
Средние величины объема потомств испытуемых деревьев
| Материнские деревья
|
Отцовские деревья
| Средние потомств
|
| 1
| 2
| 3
| 4
|
| 5
| 9
| 17
| 12
| 14
| 13
|
| 6
| 10
| 16
| 12
| 10
| 12
|
| 7
| 11
| 20
| 10
| 15
| 14
|
| 8
| 14
| 15
| 6
| 17
| 12
|
| Средние потомств
| 11
| 17
| 10
| 14
| 13 (средняя по опыту)
|
Специфическая комбинационная способность (СКС) – это термин, относящийся к средней величине потомства, полученного от скрещивания двух определенных родителей, которая отличается от величины, ожидающейся с учетом только их ОКС. Она так же может быть отрицательной или положительной. СКС всегда относится к специфическому кроссу (гибриду) и никогда – к отдельному родителю самому по себе.
СКС для кросса между деревом № 3 и деревом № 6 (кроссовое значение равно 12) может быть рассчитана следующим образом:
1. Рассчитывается ОКС для обоих родителей:
ОКС3 = - 3; ОКС6 = - 1.
2. Значения ОКС прибавляются к популяционной средней, образуя ожидаемую ценность (значение) кросса (3 х 6), базирующейся на общих комбинационных способностях.
Ожидаемая ценность = средняя опыта + ОКС3 + ОКС6 = 13 + (- 3) + (- 1) = 9.
3. Вычитаем рассчитанную ожидаемую ценность (в п. 2) от наблюдаемой ценности (значения) кросса (3 х 6). Результат и есть СКС.
СКС6 х 3 = наблюдаемое значение – ожидаемое значение = 12 – 9 = + 3.
Это означает, что кросс 6 х 3 на 3 объемных единицы лучше, чем можно было бы ожидать, исходя из значений ОКС родителей 3 и 6.
Следует подчеркнуть, что ничего нельзя сказать о пользе кроссов, основываясь только на их СКС. Поскольку СКС есть отклонение от ожидаемых значений, основанных на значениях ОКС. Кросс может иметь положительную СКС, но не быть еще хорошим по отношению к другим кроссам, как это показано на примере с родителями № 6 и № 3.
Кросс имеет СКС = + 3. Но его среднее значение, равное 12 единицам, все еще ниже среднего популяционного (13 единиц), из-за низких значений ОКС обоих использованных родителей.
Необходимо подчеркнуть здесь, что ОКС, селекционная ценность и СКС являются показателями генетической ценности родителей или кроссов только по исследуемому признаку (в данном случае объему). Например, родители могут иметь выше средние значения ОКС для объема и в то же самое время ниже средние ОКС для плотности.
Рассматривая вышеприведенный пример, следует обратить внимание еще на одну сторону вопроса. Первоначально значения ОКС и СКС определялись для инбредных линий, и методы их оценки, предложенные Г.Ф. Спрэгом и Л. А. Тейтумом в сороковых и Б. Гриффингом в середине 50-х годов, относились именно к таким инбредным линиям. Однако впоследствии эти методы применялись и к другим видам исходного материала сельскохозяйственных растений, а затем и к лесным древесным растениям (Э. Л. Березин, 1981; Я. Биргелис и др., 1981; А. М. Данченко, Г. Б. Дубинин, 1981; В. И. Долголиков, 1981; А. П. Царев, 1981; B. Zobеl, J. Talbert, 1984; С. П. Погиба, 1991).
Два типа комбинационной способности отражают различные типы взаимодействия между аллелями в генном локусе. ОКС представляет среднее значение потомства родителя, когда он скрещивается со многими другими родителями. Поэтому она отражает аддитивную генетическую ценность родителя. Это значит, что она отражает ту часть его генотипа для определенного признака, которую родитель может передать своему потомству, независимо от того, какие другие родители вовлечены в скрещивание. ОКС представляет аддитивный тип действия генов, упомянутых ранее. Родители, о которых известно, что они имеют высокую ОКС для данного признака, рассматриваются как хорошие партнеры для скрещиваний. Но они не всегда желательны. Почему? Потому, что одни из них могут иметь высокую комбинационную способность по признаку чувствительности к болезням, кривизне ствола или другим наблюдаемым характеристикам.
Поскольку ОКС позволяет предсказать аддитивный эффект, ее иногда рассматривают как зависимую часть генетической конституции дерева. Это часть комбинационной способности, которая реализуется в лесных семенных плантациях, которые состоят из многих родителей. Улучшенное значение высаженных деревьев, полученных на лесных семенных плантациях, обусловлено накоплением предпочтительных аллелей, которые оказывают аддитивный генетический эффект на фенотипы деревьев, полученных из лесных семенных плантаций.
СКС, найденная для данного кросса, отражает взаимодействие двух аллелей генного локуса, воздействующих на данный признак (доминантное действие гена) и взаимодействие между аллелями различных генных локусов, влияющих на признак, или эпистатическое действие генов. Как показано выше, эти два типа генетического воздействия обычно относятся к неаддитивным эффектам генов. СКС может быть показателем в большей части доминантного типа генного действия. Так как СКС проявляется в результате взаимодействия между специфическими аллелями, или между генными локусами, ее значение не может быть предсказано по фенотипам родителей пока не произведены скрещивания. Она не может быть использована в программах создания лесных семенных плантаций, включающих многих родителей, поскольку свободное опыление результируется во многих различных комбинациях аллелей генных локусов.
Существуют два пути использования СКС в селекционных программах. Первый – использование вегетативного размножения, чтобы производить значительные количества репродуктивного материала идентичного дереву, от которого он взят. При вегетативном размножении генотип родителей остается неизменным и СКС аллелей всех генных локусов сохраняется.
Второй путь – использование СКС для создания кроссов с целью массового производства семян от определенной комбинации родителей. Это может быть реализовано через контролируемое опыление и методом создания биклоновых семенных плантаций.
Оба этих метода, особенно метод вегетативного размножения используются в некоторых породах для производства улучшенного репродуктивного посадочного материала. Для большинства видов, однако, стоимость и технологические трудности, связанные с использованием СКС делают, делают ОКС более предпочтительной при реализации селекционных программ.
Наследуемость
Наследуемость количественного признака – это одно из важных и наиболее употребительных положений количественной генетики. Величина наследуемости выражает долю изменчивости в популяции, которая определяет генетические различия между индивидуумами.
Термин наследуемость часто заменяют термином коэффициент наследуемости, что одно и то же и означает отношение, указывающее с какой степенью родители передают свои свойства их потомству. По показателю наследуемости можно производить оценку выигрыша, который может быть получен от селекционной программы. Различают два вида наследуемости: в широком смысле (Н2) и в узком смысле (h 2). Наследуемость в широком смысле определяется как отношение общей генетической, или генотипической, изменчивости в популяции к общей фенотипической:
, (14.8)
где: σ2 G – генотипическая дисперсия;
σ2 P – фенотипическая дисперсия;
σ2 A – аддитивная дисперсия;
σ2 NA – неаддитивная дисперсия;
σ2 E – экологическая дисперсия.
В свою очередь, неаддитивная изменчивость (σ2 NA) может состоять из доминантной (σ2 D) и эпистатической (σ2 I) дисперсий.
Наследуемость в широком смысле может варьировать от 0 до 1. Низкие значения наследуемости (близкие к 0) могут встречаться, если в популяции отсутствует изменчивость, обусловленная генетическими факторами (например, при клонировании растений). Если вся изменчивость обусловлена генетическими факторами, то наследуемость будет равна 1. Наследуемость в широком смысле имеет ограниченное применение в селекции деревьев, и ее основное использование, когда обе, и аддитивная и неаддитивная изменчивости могут быть переданы от родителей потомкам полностью, например, при вегетативном размножении.
Наследуемость в узком смысле – это отношение аддитивной генетической дисперсии к общей фенотипической дисперсии:
(14.9)
Значения наследуемости в узком смысле так же колеблются от 0 (отсутствие аддитивной дисперсии) до 1 (отсутствие средовой или неаддитивной дисперсии). Наследуемость в узком смысле никогда не может превышать наследуемости в широком смысле. Если вся генетическая дисперсия вызвана аддитивной изменчивостью, то наследуемости в узком и широком смыслах равны.
Большинство оценок наследуемости, приведенных в генетической литературе по лесным древесным породам, даны для наследуемости в узком смысле, поэтому большинство современных селекционных программ направлено на улучшение ОКС и, таким образом, на использование только аддитивной части генетической дисперсии. Это, несомненно, может измениться при использовании методов вегетативного размножения или развитии экономичных методов производства специфических скрещиваний, таких, как дополнительное массовое опыление, или при создании биклоновых плантаций.
Важным, но часто игнорируемым аспектом оценок наследуемости, является их применимость только к отдельным популяциям, растущих в определенных условиях и в определенном отрезке времени. (B. Zobel, J. Talbert, 1984). Например, оценки наследуемости для групп деревьев, растущих в теплице, не будут подходящими для тех же самых деревьев, растущих в условиях открытого грунта. На высоту в теплице и в условиях открытого грунта одни и те же гены воздействуют по-разному. Даже, если оба признака будут одинаковыми, оценки h2 , полученные в теплице, будут обычно выше, чем в условиях открытого грунта, из-за более низкой экологической изменчивости в теплице. Как можно видеть из формулы h2 (14.9), изменения в компоненте экологической дисперсии (s2Е) в знаменателе будут прямо влиять на h2. Из-за влияния условий на отношение наследуемости, оценки h2 для данного признака вида в одном географическом районе, вероятно, не будут такими же, как в другом районе. Оценки наследуемости данной характеристики в популяции часто меняются с возрастом, когда внешние условия изменяются и когда генетический контроль характеристик становится другим по мере взросления дерева. Степень изменения с возрастом дискутируется, однако, наличие самого изменения мало кем оспаривается.
Для оценки наследуемости в лесной генетике используют схемы выращивания потомства от групп родителей или кроссов в одном и том же испытательном насаждении. Оценки наследуемости затем получают от соответствующих показателей потомков внутри и между родительскими деревьями. Селекционные схемы и экспериментальные планы, которые могут быть использованы для оценок дисперсий и наследуемостей представляют тему отдельного рассмотрения (Дж. У. Снедекор, 1961; С. Пирс, 1969 и др.).
Оценки наследуемости в узком смысле для высоты, плотности и формы для некоторых видов представлены в таблице (14.3). Некоторые характеристики, по данным этой таблицы, такие как плотность древесины, явно находятся под строгим генетическим контролем, независимо от вида, и являются близкими для различных условий местопроизрастания. Другие характеристики, такие как рост в высоту, находятся под меньшей степенью генетического контроля и существенно зависят от условий местопроизрастания, в которых живут деревья.
Даже в широких экспериментах с многими семьями наследуемость не может быть оценена без ошибки. Все оценки h2 должны осмысливаться как цифры, которые свидетельствуют об общей величине наследуемости. Например, наследуемость равная 0,15, не должна рассматриваться как слишком отличная от величины наследуемости в 0,20. Достоверность многих оценок наследуемости можно видеть в табл. (14.3), где наследуемость высоты черного ореха (Juglans nigra) в 8-ми летнем возрасте записана как h2 = 1,25. Ранее уже было показано, что наследуемость не может превышать единицу. Таким образом, значение h2 = 1,25 является слишком завышенным. Абсолютные оценки наследуемости необходимы для многих целей, таких как оценка выигрыша, где h2 есть неотделимая часть формулы выигрыша.
Однако, главная ценность концепции наследуемости для лесных селекционеров есть указание общей величины генетического контроля и лучшего подхода для использования в программах селекции лесных древесных пород.
Таблица 14.3