По отдельным опытам с лесными древесными породами
(по B. Zobel, J. Talbert, 1984)
| Признак
| Наследуемость
| Источник
|
|
Высота:
Pseudotsuga menziesii
Pinus taeda
Pinus taeda
Pinus elliottii
Pinus palustris, возраст 5 лет
Pinus palustris, возраст 7 лет
Liriodendron tulipifera
Juglans nigra, возраст 1 год
Juglans nigra, возраст 8 лет
Liquidambar straciflua, возраст 2 года
Liquidambar straciflua, возраст 11 лет
Плотность древесины:
Pinus taeda
Pinus taeda
Pinus sylvestris
Pinus elliottii
Eucalyptus deglupta
Eucalyptus viminalis
Форма
Pinus taeda – прямизна ствола
Pinus taeda – форма кроны
Pinus elliottii – высота очищения от сучьев
Pseudotsuga menziesii – черенкование
|
0,10-0,30
0,44
0,14-0,26
0,03-0,37
0,18
0,12
0,42-0,84
0,55
1.25
0,25
0,08
0,76-0,87
0,41
0,46-0,56
0,50
0,44
0,55
0,14-0,21
0,08-0,09
0,36-0,64
0,26
|
Cambell (1972)
Martziras and Zobel (1973)
Stonecypher et all (1973)
Barber (1964)
Snyder and Namkoong (1978)
Snyder and Namkoong (1978)
Kellison (1970)
McKleand (1978)
McKleand (1978)
Ferguson and Cooper (1977)
Ferguson and Cooper (1977)
Goggarns (1961)
Chuntanaparb (1973)
Personn (1972)
Goddard and Cole (1966)
Davidson (1972)
Otegbye and Kellison (1980)
Stonecypher et al (1973)
Stonecypher et al (1973)
Barber (1964)
Silen (1978)
|
Примечание: оценки наследуемости варьируют в зависимости от вида древесной породы, популяций внутри вида, возраста и оцениваемых признаков.
Несмотря на ее полезность, необходимо помнить, что наследуемость не является неизменной величиной, постоянной для популяции. Наследуемость сама по себе является изменчивой величиной. Поэтому, всякий раз, когда наследуемость используется для определения генетического выигрыша или стратегии селекции, оценки должны рассматриваться на вероятностном уровне, который всегда ниже 100% (B. Zobel, J. Talbert, 1984).
Методы оценки наследуемости
Для оценки наследуемости в широком смысле (H²) в лесных популяциях применяются следующие методы:
– Расчет в клоновых популяциях;
– Оценка в популяциях при выровненных экологических условиях;
– Дисперсионный анализ;
– Метод Шрикганди;
– Метод фоновых признаков В. А. Драгавцева.
Для оценки наследуемости в узком смысле (h²) используют расчет:
– По корреляции признака у одного из родителей и потомства;
– По корреляции признака у полусибсов;
– По коэффициенту регрессии;
– По внутриклассовой корреляции;
– Способом анализа полных сибсов (сиб-анализ).
В качестве примера для оценки наследуемости в широком смысле приведем расчет H² в клоновых популяциях. В лесном хозяйстве к таковым относятся плантации вегетативного происхождения (прививочные, черенковые и др.), порослевые насаждения лиственных пород, корнеотпрысковые древостои и др.
Основой расчета H² в таких насаждениях является генотипическая однородность деревьев внутри клона и генотипическая разнородность клонов между собой. В этом случае вся видимая (фенотипическая) изменчивость между деревьями одного клона будет экологической, то есть:
σ ² P = σ ² E (14.10)
Изменчивость между всеми деревьями в популяции, выраженная фенотипической дисперсией, обусловлена различиями между генотипами клонов и неоднородностью экологических условий, то есть:
σ ² P = σ2 G + σ ² E (14.11)
Значения изменчивости между всеми деревьями всей популяции и между деревьями внутри каждого клона вычисляются по фактическим наблюдениям. Тогда:
σ2 G = σ ² P - σ ² E (14.12)
То есть, генотипическая дисперсия в клоновых популяциях равна фенотипической за минусом средней величины экологической дисперсии внутри клонов. А отсюда коэффициент наследуемости в широком смысле может быть определен по формуле (14.8), приведенной выше.
Определение наследуемости в узком смысле несколько сложнее, поскольку сложнее определить аддитивную составляющую дисперсии. Поэтому предлагаемые перечисленные выше методы направлены на то, чтобы обойти это препятствие. Для примера рассмотрим метод определения коэффициента наследуемости в узком смысле (h²) с использованием коэффициентов пути С. Райта.
Метод Райта основан на сопоставлении наблюдаемых (эмпирических) значений коэффициентов корреляции между фенотипами родственных особей с соответствующими коэффициентами корреляции между их генотипами.
Связь между фенотипами родителей и потомков может быть с некоторым упрощением изображена в виде следующей схемы (рис. 14.3), где
– 0,5 – коэффициент корреляции между генотипами родителей и потомков
– h = rgp – коэффициент корреляции, характеризующий степень связи между генотипическими и фенотипическими различиями особей, или коэффициент корреляции между фенотипом p и генотипом g (коэффициент пути от генотипа к фенотипу).
– r – коэффициент корреляции между фенотипами родителей и потомков.
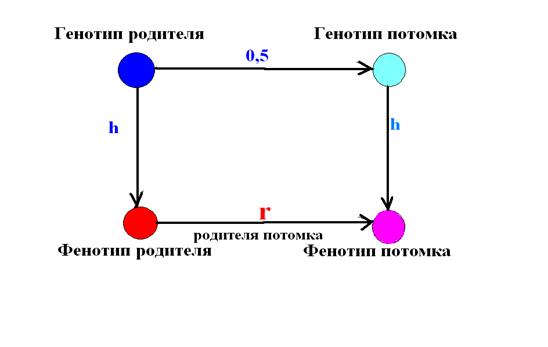
Рис. 14.3. Примерная схема коэффициентов пути С. Райта.
При использовании метода оценки наследуемости в узком смысле, основанном на путевых коэффициента С. Райта, вводятся некоторые допущения и значения для корреляции.
Так, корреляция между генотипом родителя и потомка rpo = 0,5 проявляется при следующих допущениях:
– признаки наследуются аддитивно;
– популяция находится в состоянии равновесия;
– в популяции происходит свободное скрещивание.
При этих допущениях коэффициент корреляции равен:
– между генотипами полных сибсов 0,5;
– между генотипами дедов и внуков 0,25;
– между генотипами полусибсов 0,25.
Приведенная схема показывает, что величина корреляции между фенотипами родителей и потомков rpo зависит от:
– величины корреляции между фенотипом родителя и его генотипом (h),
– корреляции между генотипами родителей и потомков (r=0,5),
– величины корреляции между фенотипом потомка и его генотипом (h).
Применяя разработанный С. Райтом метод коэффициентов пути (path coefficients), находим, что коэффициент корреляции rpo равен произведению всех коэффициентов цепи связей, соединяющей фенотип родителя с фенотипом потомка, т.е
rpo = 0,5 h ², (14.13)
откуда
h ² = 2 rpo (14.14).
То есть, наследуемость в узком смысле для каждого потомка от одного родителя равна удвоенному фенотипическому коэффициенту между ними.
Здесь разобраны простые случаи только двух методов. Для разбора более сложных случаев и практики применения других методов следует использовать специальную литературу (П. Р. Леппер, З. С. Никоро, 1966; П. Ф. Рокицкий, 1973; Н. А. Плохинский, 1964; М. М. Котов, 1997; В. А. Драгавцев, 1973; и др.).






