Инбридинг означает скрещивание (спаривание) особей, связанных друг с другом родством. Крайний случай - это самоопыление или опыление дерева собственной пыльцой. Однако имеются различные градации инбридинга, начиная с этой крайности: скрещивание между братом и сестрой, между двоюродными братьями и сестрами, между членами небольших изолированных популяций. Степень родства в популяции зависит от ее размера. В популяции двуполых организмов каждая особь имеет двух родителей, четырех прародителей, восемь прапрародителей и т.д., так что t поколений назад у нее получится 2 t предков. Понятно, что даже для небольшого числа поколений t число всех различных предков, исключающее инбридинг, будет больше, чем любая реальная популяция.
Следовательно, особи любой пары должны быть связанными одним или более общими предками в более или менее отдаленном прошлом. При этом чем меньше размер популяции в предшествовавших поколениях, тем менее отдалены эти предки и тем большее их число. Таким образом, все родительские пары связаны в малой популяции более тесным родством, чем в большой. Вследствие этого свойства малых популяций можно рассматривать как последствия инбридинга.
Эти свойства малых популяций у древесных растений могут проявляться:
1) в единообразии внутри и изменчивости между инбредными линиями,
2) потере мощности роста,
3) снижении урожая семян,
4) ослаблении или отсутствии адаптации и др.
Проиллюстрируем это примерами, приводимыми Д. В. Райтом (1978).
Единообразие внутри и изменчивость между инбредными популяциями. Допустим, что гетерозиготное (имеет разные аллели Аа) по какому-то отдельному локусу дерево самоопылено. Генетический состав его потомства будет: 1 АА:2 Аа:1 аа. То есть, 1/4 потомства будет гомозиготна по АА и 1/4 гомозиготна по аа, или в целом, половина потомства будет гомозиготна по этому отдельному локусу. В соответствии с этим, самоопыленное гомозиготное по АА дерево даст гомозиготное потомство АА, а гомозиготное по аа дерево даст потомство, гомозиготное по аа. У каждого поколения число гетерозигот будет уменьшаться наполовину.
Таким образом, начав с дерева с генотипом АаВbСcDdEe, можно путем повторного самоопыления закончить чистыми линиями с генотипами AABBCCDDEE, aabbccddee, AABBccDDee и т.д. Каждая из этих линий однородна внутри себя, но сами линии могут значительно различаться.
То же самое может произойти, если мы имеем дело не с самоопылением, а с малой популяцией.
Потеря мощности роста. Дикие популяции деревьев обычно несут большой “генетический груз”, т.е. они гетерозиготны по большому числу вредных рецессивных генов. Эти гены не приносят вреда потому, что каждое дерево обычно опыляется другим, которое не обладает такими же рецессивными генами. Таким образом, у нормальных сеянцев вредные рецессивные гены не проявляются. Если же дерево с генотипом AaBbCc, самоопылилось, то часть потомства будет гомозиготно по аа, часть - по bb и часть - по сс.
В случае, если родительское дерево содержит 100 вредных рецессивных генов в разных локусах, его потомство будет гомозиготно в отношении вредных рецессивов в среднем в 25% локусов. Даже если отдельные гены малоэффективны, действие их на скорость роста может быть значительным. По данным лесной службы США в Райнлендере (Висконсин) средняя величина депрессии инбридинга в питомнике ели канадской достигала 40-50%.
Однако, как уже отмечалось ранее, результаты самоопыления могут быть разными. Так в опытах Е. С. Франклина с сосной ладанной (Pinus taeda) в Северной Каролине США получены следующие результаты. Из 132 самоопыленных деревьев четыре дали такое же количество семян и более мощное потомство от самоопыления, как и от скрещивания с другими деревьями. Остальные 128 показали небольшое снижение урожая семян (некоторые вообще не дали семян при самоопылении), а также меньшую мощность потомства. В экспериментах с другими видами сеянцы от самоопыления зачастую росли со скоростью, почти в два раза меньшей по сравнению с нормальными.
Ю. Н. Исаков (1999) на основании изучения изменчивости репродуктивных признаков у сосны обыкновенной (Pinus sylvestris) при самоопылении и свободном опылении определил индивидуальную изменчивость деревьев по уровню самофертильности. Им выделено 5 групп растений: 1) высокосамостерильные, 2) самостерильные, 3) частичносамофертильные, 4) самофертильные и 5) высокосамофертильные.
Изучение уровня самофертильности у одних и тех же 40-50 деревьев в течение 6 лет показало, что он варьирует в зависимости от условий среды, и в разные годы имеет разную величину.
Между уровнем самофертильности деревьев и степенью депрессии роста потомства после самоопыления выявлена отрицательная зависимость.
Как показали другие данные, снижение мощности, или “инбредая депрессия” не является постоянным следствием инбридинга. Томаты, пшеница и многие другие травянистые растения имеют структуру цветка, способствующую самоопылению. Когда-то, возможно, они и могли содержать вредные рецессивные гены, но тысячи поколений самоопыления могли их элиминировать. Поэтому современные растения томатов и пшеницы образуют одинаково мощное потомство и при самоопылении, и при перекрестном опылении.
В умеренной зоне большинство деревьев имеют механизмы, которые препятствуют самоопылению. В результате близкородственный инбридинг не снижает постоянно скорость роста. Это объясняется тем, что снижение мощности вызывается скорее аккумуляцией вредных рецессивных генов, чем самим самоопылением. Деревья, не имеющие таких рецессивных генов, могут самоопыляться без вредных последствий. То есть, самоопыление обычно вредно, но может быть и полезным, как это показали опыты Ю. Н. Исакова.
Снижение урожая семян. Вредные рецессивные гены могут действовать в период развития семян. В таких случаях оплодотворение собственной пыльцой может произойти, но оплодотворенный зародыш может абортировать на определенной стадии развития, что приводит к образованию пустого семени. Это является возможным объяснением снижением урожая семян в некоторых опытах.
Однако, для некоторых покрытосеменных, которые несут аллели самостерильности S1, S2, S3, S4 и др., может быть и другое объяснение. Эти аллели действуют так, что пыльца S1 и S2 не может прорасти на рыльце растения с генотипом S1S2, а пыльца S1 или S3 не может прорасти на рыльце растения с генотипом S1S3.
Результаты опытов по самоопылению древесных растений пока незначительны, возможно, из-за небольших масштабов некоторых экспериментов. В целом, можно выделить три группы растений по семеношению при самоопылении (Д. В. Райт, 1978):
1.Урожай семян очень низкий, потомство обычно слаборослое: Alnus glutinosa, Castanea molissima; Eucaliptus alba и др.; Larix decidua и leptolepis; Liriodendron tulipifera; Picea jezoensis и sitchensis; Pinus resinosa, strobus, sylvestris, thunbergiana, rigida; Pseudotsuga menziesii; Ulmus am e ricana, parvifolia.
2. Образуется много жизнеспособных семян: Alnus incana, Eucaliptus, Caesia и др.
3. Варьирование от незначительного до большего количества жизнеспособных сеянцев, потомство от слаборослого до мощного:
Acacia decurrenus; Caragana arborescens; Eucaliptus bicostata; Picea abies, omorica; Pinus monticola, taeda.
К последней группе можно отнести и некоторые экотипы Pinus sylvestris.
Отсутствие или снижение адаптации. Предположим, что дерево гетерозиготно по N локусам. У потомства от самоопыления будет 1/2 N или 50% этих локусов. Даже если потомки останутся в живых, многие из них будут гомозиготны по вредным генам. Следовательно, выживающая самоопыленная линия не может быть хорошо адаптирована к условиям данной среды.
При инбридинге в малых популяциях имеет место некоторая случайная фиксация генов, которая может привести к образованию неадаптированных к данной среде организмов. Примером может служить часть южных видов ели Северной Америки, некоторые из которых находятся на грани вымирания.
13.2. Измерение степени инбридинга
Степень инбридинга определяют разными способами. (П. Ф. Рокицкий,1978; Д. С. Фолконер, 1985; М. Lynch, B. Walsh, 1997). Наиболее простую концепцию излагает Д. В. Райт (1978), которая здесь и приводится.
Самоопыление является особым случаем инбридинга, при котором N =1 (N- число деревьев, входящих в перекрестноопыляющуюся популяцию). При самоопылении гетерозиготность с каждой генерацией уменьшается на 1/2. Таким образом, самоопыляющаяся линия сохраняет 1/2 исходной изменчивости после первой генерации, 1/4 - после второй, 1/8 - после третьей и т.д.
Для измерения величины инбридинга используют коэффициент инбридинга F, если N =1 и более. Коэффициент инбридинга - это вероятность того, что два гена в данном локусе идентичны по происхождению.
Коэффициент инбридинга особи выражает степень родства между ее родителями. Если родители любого поколения скрещиваются случайно, то тогда коэффициент инбридинга потомств соответствует вероятности того, что две гаметы, взятые наудачу из родительского поколения, несут идентичные по происхождению гены по данному локусу. Это будет средний коэффициент инбридинга всех потомков. Особи из разных семей будут иметь различные коэффициенты инбридинга, так как при случайном скрещивании одни пары родителей будут состоять в более тесном родстве, чем другие пары. Однако в качестве меры процесса важен именно средний коэффициент инбридинга, обозначенный как F.
F - есть потеря степени гетерозиготности, или напротив увеличение степени гомозиготности. Для выражения изменения в количестве инбридинга на одно поколение применяют обозначение D F.
Существуют две основные формулы для вычисления D F. Наиболее простая:
D F = F1 = 1/2 Nобщ (13.1)
Эту формулу применяют, если N - общее число деревьев в селектируемой популяции и если каждое дерево используется как отцовское и материнское.
Другая формула, применяемая при скрещивании разных количеств мужских N ♂ и женских N ♀ деревьев, имеет вид:
D F = F1 = (N ♂ + N ♀)/8(N ♂ N ♀) (13.2)
Эти формулы идентичны, если N ♂ = N ♀ и если N общ = 2 N ♂ = 2 N ♀
Вывод данных формул можно найти в книге П. Ф. Рокицкого (1978). Их идентичность видна из следующего простого примера. Допустим, в некоей группе имеется 20 однодомных деревьев. Тогда, используя формулу 13.1, получим:
D F = F1 = 1/2 Nобщ = 1/2·20 = 1/40.
Если такая же по численности группа, состоит из двудомных деревьев, и половина из них представляет женские экземпляры, другая половина мужские, то в соответствии с формулой 13.2 получим:
D F = F1 = (N ♂ + N ♀)/8(N ♂ N ♀) = (10 + 10)/8(10·10) = 20/800 = 1/40.
То есть, как видно из приведенных расчетов, результат является одинаковым.
Чтобы узнать степень гетерозиготности необходимо от 1 отнять величину коэффициента инбридинга, т. е. в данном случае гетерозиготность (H) за одно поколение уменьшится и будет равна:
H = 1 – F = 1 – 1/40 = 39/40 = 0,975
Кроме численных методов имеется простой метод, позволяющий определять коэффициент инбридинга для любого организма с известной родословной - метод анализа путей (П. Ф. Рокицкий,1978; Ф. Айала, Дж. Кайгер, 1988; С. И. Малецкий, 1995; М. Lynch, B. Walsh, 1997; И. Ф. Жимулев, 2003).
На рис. 13.1 и 13.2. представлены родословные, которые иллюстрируют этот подход. На рис. 13.1 изображена схема скрещивания родных братьев и сестер. Каждая стрелка соответствует передаче следующему поколению одной гаметы. Пусть A и B – не состоящие в родстве родители, из гамет которых образуются зиготы C и D. Зигота E возникает при слиянии гамет от сибсов C и D.

Рис. 13.1. Родословная потомства от скрещивания между братом и сестрой (Ф. Айала, Дж. Кайгер, 1988. Т. 3).
Поскольку A и B не состояли в родстве, считается, что их аллели в определенном локусе не идентичны по происхождению. Их можно обозначить у особи A как a 1 a 2, а у особи B – a 3 a 4 (индексы указывают, что аллели не являются идентичными по происхождению, но могут быть одинаковыми по структуре). Вероятности появления разных типов потомков от скрещивания А х В составляют ¼ (а1а3), ¼ (а1а4), ¼ (а2а3), и ¼ (а2а4). Вероятность появления в потомстве от скрещивания сибсов особей, гомозиготных по какому-либо одному аллелю, т.е. a 1 a 1 или a 2 a 2, или a 3 a 3 , или a 4 a 4 равна ¼.
Руководствуясь родословной, приведенной в рис. 13.1, можно рассчитать вероятность появления гомозиготных особей у потомка Е. Родитель А производит гаметы двух типов a 1 и a 2, каждую с вероятностью ½. Следовательно, вероятность того, что С получит от А a 1, равна ½ и вероятность того, что С передаст этот аллель (если он его имеет) своему потомку Е, также равна ½. Таким образом, вероятность того, что Е получит аллель a 1 от А через С, равна ½ х ½ = ¼. Вероятность того, что от А аллель a 1 перейдет к D, а от D к Е, также равна ½ х ½ = ¼. В результате Е получит аллель a 1 с вероятностью ¼ от С и с вероятностью ¼ от D. И как результат, вероятность того, что Е получит аллель a 1 от обоих родителей С и D, равна, ¼ х ¼ = 1/16. Этот подход можно применить и относительно любого другого аллеля (a 2, a 3, или a 4).
На основании вышеприведенной логики рассуждений разработан метод анализа путей, или путевой анализ, позволяющий определять коэффициент инбридинга для любого организма с известной родословной.
На примере родословной особи K (рис. 13.2) иллюстрируется сущность путевого анализа. Он основан на подсчете числа стрелок в родословной, образующих замкнутые циклы. На рис. 13.2 видны два цикла: K-J-G-A-D-H-K и K-J-G-C-B- D -H-K. Поскольку K появляются дважды в каждом цикле, число этапов надо сократить на единицу. Если циклы записывать не в строчку, а в виде кольца, то обозначение K появлялось бы один раз и не пришлось бы отнимать единицу от числа стрелок. Коэффициент инбридинга равен сумме слагаемых, каждое из которых определяется числом этапов в соответствующем пути и равно (1/2) n, где n равно числу этапов минус единица (или просто числу этапов, если, если рассматриваемая особь появляется в каждом пути только один раз).
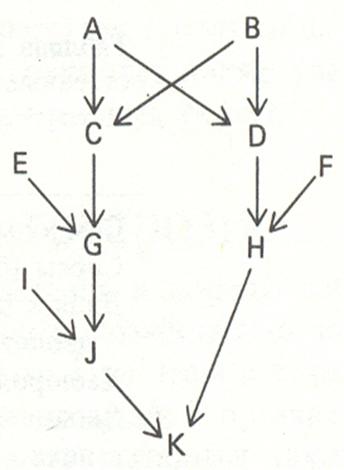
Рис. 13.2. Родословная потомства от скрещивания между двоюродными дядей (теткой) и племянницей (племянником) - (Ф. Айала, Дж. Кайгер, 1988. Т. 3).
Вероятность передачи гена от родителя потомку, как уже отмечалось ранее, всегда равна 1/2, поэтому, возведя 1/2 в степень (число этапов минус единица) и сложив полученные цифры для всех выявленных циклов, получаем искомый результат - коэффициент инбридинга. В рассматриваемом примере коэффициент инбридинга особи K равен (1/2)6 + (1/2)6 = 1/32, то есть вероятность того, что особь K гомозиготна по одному из четырех прародительских генов (a1, a2, a3, a4), равна 1/32, или 3,125 %.
При разных типах спаривания в зависимости от степени связи между родительскими формами будут разные коэффициенты инбридинга. При самооплодотворении шанс на то, что обе гаметы получают одинаковые аллели, равен 0,5. Значит, F для потомков в этом случае равен 0,5. F для потомков от спаривания полных сибсов - 0,25 и т.д.
Значения F при спаривании особей, находящихся в разной степени родства, показаны в табл. 13.1. В данном случае речь идет об оценке коэффициентов инбридинга потомков при однократном спаривании.
Таблица 13.1






