|
Индивидуумы
|
1
|
2
|
3
|
|
1
|
0,5
|
0,25
|
0
|
|
2
|
0,25
|
0,5
|
0
|
|
3
|
0
|
0
|
1
|
Обозначим оценку некоторого спаривания f2,1 = 0,25.
Поскольку ковариационная матрица симметричная, то f 2,1 = f 1,2 .
Значения показателей по диагонали (самокоанцестри) появляются только однажды.
Коэффициент родства часто представляется в такой матрице (числитель матрицы). В отсутствии инбридинга эти оценки в два раза больше.
Таким образом, коанцестри - это вероятности, и таким образом 0 # f # 1.
Некоторые примеры значений коанцестри:
Родство особей Коанцестри
Отсутствует 0
Полусибсы 0,125
Полные сибсы 0,25
Родители-потомки 0,25.
Групповое коанцестри
Если все гомологичные гены поместить в один большой пул и выбрать наугад два из них (с замещением), то вероятность того, что они являются идентичными по происхождению, определяется как групповое коанцестри ( ). Этот термин был предложен Кокерхэмом (Dag Lindgren, 1999).
). Этот термин был предложен Кокерхэмом (Dag Lindgren, 1999).
Групповое коанцестри равняется среднему числу всех N2 значений коанцестри всех комбинаций N индивидуумов в популяции (или среднее число всех 4 N2 комбинаций индивидуальных генов). Возможно, также определять групповое коанцестри как среднее число. Преимущество вероятностного определения проявляется в более сложных ситуациях.
Сформируем, или получим, среднее число из матрицы коанцестри(табл. 12.3).
Сумма из 9 оценок в матрице = 2,5. Среднее число = групповому коанцестри = 2,5/9 = 0,278. То есть, групповое коанцестри здесь - это среднее число.
Следует обратить внимание на то, что показатели самокоанцестри появляются однажды, в то время как другие показатели коанцестри появляются дважды (реципрокно). Если все индивидуумы в популяции имеют тот же самый тип, достаточно вычислить N показателей коанцестри для одного отдельного индивидуума. Самокоанцестри - это групповое коанцестри для популяции с одним единственным членом.
Все члены в семействе полных сибсов имеют одинаковый показатель коанцестри. Таким образом, достаточно построить матрицу коанцестри для семейства полных сибсов, чтобы иметь представление о некоторых генетических параметрах исследуемой совокупности деревьев. Групповое коанцестри зависит от родства, не зависимо от того, как гены объединены в гаметы. Брат одинаково связан с его братом, как и с его сестрой, несмотря на то, что его гаметы способны объединиться только с таковыми его сестры.
Если в популяции известны инбридинг и коанцестри, групповое коанцестри может быть рассчитано. Формула для группового коанцестри в этом случае может быть следующей:
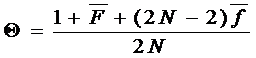 (12.4)
(12.4)
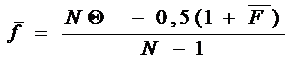 (12.5)
(12.5)
Здесь 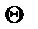 - групповое коанцестри; N - индивидуумы;
- групповое коанцестри; N - индивидуумы;
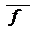 - среднее значение коанцестри для различных индивидуумов;
- среднее значение коанцестри для различных индивидуумов;  - среднее значение инбридинга.
- среднее значение инбридинга.
Изменения группового коанцестри при изменении поколения можно вычислить ретроспективно, если известна родословная, связывающая данную популяцию с основателями.
Будущее групповое коанцестри может быть рассчитано, если имеется представление о будущих родословных.
Для других случаев прогноз возможен также, однако, это более сложный процесс.
Связь между поколениями осуществляется через гаметы. Генный пул потомства идентичный генному пулу успешных гамет родителей.
Рассмотрим пару генов, которые могут быть эквивалентно расценены как в зиготах потомства, так и в успешных родительских гаметах. Пара генов может быть идентичны по происхождению, поскольку они копии того же самого гена в родительской популяции. Это может случаться, если родитель имеет больше чем одного потомка.
Пара генов может происходить из однородных генов той же самой родительской зиготы в родительской популяции. Если был инбридинг, рассматриваемые гены могут быть идентичны по происхождению.
Сибсы - это те особи, которые делят одного родителя, и получают коанцестри равное (1 + Fp) / 2. Если рассматриваемая пара генов происходит от различных родителей, коанцестри будет равным fp. Индекс p при показателях инбридинга(F) и коанцестри (f) означает, что данные показатели относятся к родительским поколениям.
Идентичность по происхождению может обеспечиваться следующими механизмами:
1. Тот же самый ген в текущем поколении отбирается дважды;
2. Гены являются копиями того же самого гена в родительском поколении;
3. Гены происходят от гомологичных генов в одном и том же инбредном родителе;
4. Гены приходят от различных, но связанных родством, родителей.
Групповое коанцестри обеспечивает вероятность того, что два гена в популяции являются идентичными по происхождению.
Разнообразие означает, что мы имеем дело с различными объектами. Разнообразие генов означает, что гены различны.
Очевидно, что групповое коанцестри может быть оценено как 1 (единица), если существует полная вероятность того, что гены не являются идентичными, то есть они разнообразны. Для оценки разнообразия генов в какой-либо популяции может быть использована формула:
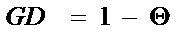 , (12.6)
, (12.6)
где GD - мера разнообразия генов, а групповое коанцестри () - мера потерянного разнообразия генов в рассматриваемой популяции.
Этот подход предполагает, что все гены в индивидуумах рассматриваемых популяций уникальны. Мера, основанная на групповом коанцестри, относится к рассматриваемой популяции. Генное разнообразие GD - является ожидаемым средним значением гетерозиготности.
Для селекции лесных древесных пород дикий лес часто представляет хорошую основу. Разнообразие генов дикого леса равно 1, и групповое коанцестри может дать представление о доле или проценте потерянного разнообразия генов.
Если контролировать групповое коанцестри при различных приемах селекции лесных древесных пород, то можно сказать, как много было потеряно разнообразия генов по сравнению с диким лесом.
Понятие об уставном числе
Уставное число (status number, Ns) - это половина обратной величины группового коанцестри. Или, что является эквивалентным, - это половина обратной величины вероятности того, что два гена, взятые случайно являются идентичными по происхождению. Этот показатель ввел в научный обиход шведский ученый Даг Линдгрен.
Привлекательная особенность уставного числа заключается в том, что оно отражает списочное число (перечень) N неродственных и неинбредных индивидуумов в популяции. Уставное число говорит о том, что вероятность отобрать два идентичных по происхождению гена есть такая же, как если бы это было число неродственных неинбредных индивидуумов, составляющих генный пул. Поэтому его можно называть эффективным числом (D. Lindgren, 1999).
Уставное число - интуитивно более привлекательный путь представления группового коанцестри, поскольку это связано с известной концепцией размера популяции.
Отношение уставного числа и списочного числа называется относительным уставным числом (Nr):
Nr = Ns/N. (12.7)
Основные свойства уставного числа следующие:
- Оно никогда не может быть выше, чем списочное число (N);
- Оно никогда не может быть более низким, чем 0,5 (оценка гаметы);
- Оно рассматривает родство и инбридинг;
- Не имеет значения, если члены "популяции" принадлежат тому же самому поколению или той же самой "подпопуляции". Уставное число может быть рассчитано для любой гипотетической популяции (с известной родословной).
- Это - не то же самое, что инбридинг, но оно тесно связано с инбридингом, поскольку инбридинг, наступая после равного количества всех возможных спариваний, будет равен 0,5/ Ns. После случайного спаривания это является ожидаемой величиной. Но инбридинг может быть равным 1, в то время как уставное число - 0,5 # Ns # N /2. Инбридинг также может быть равным 0, в то время как уставное число - 1 # Ns # N.
- Если имеется ряд дискретных поколений с различным списочным числом, Ns не может превышать минимум N в предшествующих поколениях.
- Для уставного числа не имеет значения род членов популяции.
- Уставное число после смены поколения зависит только от численности потомства для каждого родителя. Оно не зависит от того, как родители спаривались. Уставное число описывает генный пул, а не то, как он образован.
- Уставное число обычно снижается при смене поколений, но оно может и повышаться, поскольку первоначальные геномы могут иметь более равное представительство после изменения поколения, чем прежде.
- Уставное число может быть рассчитано для любой популяции с известной родословной, связывающей ее с основателями с известным коанцестри и инбридингом (подобно вычислениям инбридинга). Для этого Дагом Линдгреном с сотрудниками разработаны специальные алгоритмы (D. Lindgren at al, 1997; D. Lindgren, 1999).
С помощью уставного числа и группового коанцестри можно измерять дисперсию гена. Согласно Кокерхэму (1969, цит. D. Lindgren, 1999) варианса частоты гена (p), или среднего значения встречаемости гена есть:
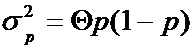 , (12.8)
, (12.8)
или
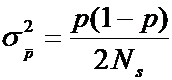 . (12.9)
. (12.9)
Это - биномиальное выражение вариансы для частоты гена в популяции с Ns неинбредных и неродственных членов.
Таким образом, уставное число - это количество неродственных и неинбредных деревьев, отобранных из исследуемой популяции, которые испытали тот же самый дрейф, как и накопленный дрейф данной популяции.
Концепция уставного числа, по мнению Дуга Линдгрена, для лесных селекционеров более полезна, чем для генетиков и селекционеров, работающих с другими объектами. Это связано с тем, что лесные селекционеры ближе к основателям популяций и вообще имеют дело во многих случаях с диким лесом. Они имеют дело с немногим числом поколений, имеющих специфические смены между собой. Кроме того, лесные селекционеры могут осуществлять контроль над популяциями в течение длительного времени.
В заключение необходимо отметить, что то краткое изложение понятий и терминов, которые рассмотрены в настоящей и предыдущей главе, дает только первое представление о количественной генетике. Более полное знание об этом предмете можно почерпнуть как уже из цитировавшихся книг зарубежных исследователей Namcoong, 1981; Д. С. Фолконера, 1985; J.; D. S. Falconer, 1989; D. S. Falconer, T. F. S. Mackay, 1996; М.Lynch, B.Walsh, 1997; Dag Lindgren at al., 1997, a, b, c; Dag Lindgren, 1999 и др.; так и из работ отечественных авторов: П. Ф. Рокицкого, 1978; В. А. Драгавцева, 1979; Н. В. Глотова и др., 1982; Л. А. Животовского, 1991, и др.
Вопросы для самопроверки
1. Какие типы родства являются важными для лесной генетики?
2. Приведите формулу степени сходства для полных сибсов, выраженную через коэффициент регрессии от потомков к родителям.
3. Что такое ковариация? Чему равна ковариация:
а) потомков и одного родителя;
б) потомков и среднего из значений родителей;
в) полусибсов;
г) полных сибсов?
4. Чему равна регрессия (b):
а) потомков на одного родителя;
в) потомков на родительское среднее?
5. Чему равна корреляция (t):
а) между полусибсами;
б) между полными сибсами?
6. Что такое коанцестри? Дайте определение понятий идентичный по происхождению, основатель популяции, самокоанцестри, генный пул. Приведите примеры значений коанцестри при различной степени родства особей.
7. Что такое групповое коанцестри? Как оно рассчитывается? Приведите формулы расчета группового коанцестри и среднего значения коанцестри для различных индивидуумов.
8. Как осуществляется связь между поколениями? Какими механизмами обеспечивается идентичность генов по происхождению?
9. Покажите связь между групповым коанцестри и разнообразием генов. Приведите формулу для определения генетического разнообразия.
10. Что такое уставное число? Назовите его основные свойства.
11. Приведите формулы вычисления дисперсии частоты гена с помощью группового коанцестри и уставного числа.
Глава 13.
ИНБРИДИНГ






