

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни...

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Когда производится ограждение поезда, остановившегося на перегоне: Во всех случаях немедленно должно быть ограждено место препятствия для движения поездов на смежном пути двухпутного...
Марксистская теория происхождения государства: По мнению Маркса и Энгельса, в основе развития общества, происходящих в нем изменений лежит...
Интересное:
Принципы управления денежными потоками: одним из методов контроля за состоянием денежной наличности является...
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Сходство (по англ. – resemblance) между родственниками является одним из наиболее значительных явлений, которое используется в количественной генетике. Зная его, можно рассчитать количество аддитивной генетической дисперсии, что в свою очередь позволяет выбрать наилучший метод селекции.
В таблице 11.9 и формулах (11.1) и (11.16) представлено разложение фенотипической дисперсии на компоненты, относящиеся к действию разных причин. Эти компоненты Д. С. Фолконер (1985) предложил называть казуальными, или причинными и обозначать символом V.
Измерение степени сходства между родственниками опирается на иной способ разложения фенотипической дисперсии – на компоненты, соответствующие распределению особей по семьям. Эти компоненты можно вычислить непосредственно по фенотипическим значениям; называют их наблюдаемыми компонентами и обозначают символом σ 2.
Важными для лесной генетики являются 4 типа родства:
1) потомки и один родитель;
2) потомки и родительское среднее (из значений матери и отца);
3) полусибсы (потомки, у которых один известный родитель, другой – нет);
4) полные сибсы (потомки, у которых оба родителя известны).
Для примера рассмотрим распределение особей по семьям, состоящим из полных сибсов. Дисперсию в данном случае можно разложить на 2 компоненты: межгрупповую (или дисперсию групп) и внутригрупповую дисперсию. Межгрупповая компонента соответствует дисперсии “истинных” групповых средних вокруг популяционного среднего, а внутригрупповая – дисперсии вокруг истинных средних своих групп. Сходство между полными сибсами можно рассматривать и как подобие особей одной и той же группы, и как различие между особями разных групп. Чем больше будет подобие внутри групп, тем больше будет различие между группами. Таким образом, степень сходства можно определить, выразив межгрупповую дисперсию как долю общей дисперсии, заданную соотношением:
|
|
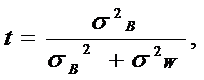 |
где: σ2В - межгрупповая компонента, а σ2W – внутригрупповая компонента.
Межгрупповая дисперсия отражает ту долю изменчивости, которая является общей для членов одной и той же группы, и представляет ковариацию членов групп. Отметим, что ковариация (или коварианса)– это сопряженная изменчивость двух признаков.
Определение ковариации является первым шагом для установления коэффициента корреляции. Корреляция - это тенденция двух мер на разных индивидуумах, или двух разных мер на одном и том же объекте, варьировать параллельно. Например, высота и диаметр одного итого же дерева или длина руки и ноги у одного и того же человека, или его высота и вес. Можно ожидать, что в первых двух случаях, чем больше первый показатель (высота дерева и длина руки человека), тем больше будет и второй (диаметр и длина ноги). Меньше оснований ожидать, что более высокие люди являются и более тяжелыми. Для количественной оценки этих ожиданий используется коэффициент корреляции.
Коэффициент корреляции - это отвлеченное число, измеряющее сопряженное варьирование (или ковариацию). Если размер каждого из сопоставляемых признаков варьирует, то их корреляция является мерой такой величины, которая входит в оба эти варьирования. Для вычисления корреляции следует учитывать и среднюю величину, и дисперсию изучаемой характеристики. Сначала определяется коварианса путем умножения отклонения точки наблюдения по оси X от средней ¯X и её же отклонение по оси Y от средней ¯Y.
В интернетовском курсе лекций по количественной генетике (Biology B 31 - Quantitative Genetics 2, 1998) приводится следующий пример вычисления ковариансы. Рассматривается группа отцов следующего роста: 1,4 м, 1,5 м, 1,6 м, 1,7 м, 1,8 м. Средняя высота группы - 1,6 м. Дисперсия высот равна:
|
|
s2 = [(1,4 - 1,6)2 + (1,5 - 1,6)2 + (1,6 - 1,6)2 + (1,7 - 1,6)2 + (1,8 - 1,6)2]/ 5 = 0,02
Среднеквадратическое отклонение равно:
s = √ 0,02 = 0,1414.
Каждый из этих отцов имеет сына следующего роста: 1,6м, 1,7 м, 1,6 м, 1,9 м, 2,2 м.
Исходя из этих данных, была рассчитана коварианса (табл.12.1).
Таблица 12.1
Вспомогательная таблица для расчета ковариансы отцы-сыновья
| Номера пар | Высоты отцов | Высоты сыновей | Коварианса |
| 1 2 3 4 5 | 1,4 1,5 1,6 1,7 1,8 | 1,6 1,7 1,6 1,9 2,2 | (-0,2) х (-0.2) = 0.04 (-0,1) х (-0.1) = 0.01 0 х (-0,2) = 0 0,1 х 0,1 = 0,01 0,2 х 0,4 = 0,08 |
| Средняя | 1,6 | 1,8 | Коварианса (Covpo) = 0,14* |
* Нижние индексы означают: o – offspring (англ.) – потомок, отпрыск; p – parent (англ.) - отец, мать или родители (мн. ч.).
Данный пример демонстрирует путь расчета ковариансы. Из него видно, что высокие отцы имеют высоких сыновей. То есть, это пример положительной корреляции. Коэффициенты корреляции могут варьировать от 0 до 1 и быть положительными или отрицательными.
Для расчета коэффициента корреляции (r) необходимо разделить ковариансу на меру варьирования отцов (σp) и сыновей (σo):
r = Covpo / σp x σo. (12.2)
В силу менделевских законов между родственниками существуют генетические корреляции. То есть, у отца и сына половина генов являются общими; у братьев и сестер также общими являются половины их генов; двоюродные братья и сестры имеют только по 1/8 общих генов; у однояйцовых близнецов - все гены общие.
В случае, когда речь идет о сходстве потомков с родителями, наблюдения группируются в пары, а не в группы; один из родителей или среднее двух родителей ставятся в пару к одному из потомков или среднему из нескольких потомков. Ковариация потомков с родителями Covop вычисляется из суммы попарных произведений, а степень сходства выражается как регрессия от потомков к родителям по следующей формуле:
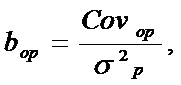 |
где: Covop – ковариация потомков с родителями, а σ2р – дисперсия признака у родителей.
|
|
Фенотипическое сходство между родственниками различной степени родства представлено в табл. 12.2.
Таблица 12.2
Фенотипическое сходство между родственниками*
(По Д.С. Фолконеру, 1985)
| Родственники | Ковариация | Регрессия (b) и корреляция (t) |
| Потомки и один родитель | ½ VA = 1/2σ2A | b = ½ VA /VP = 1/2σ2A / σ2P |
| Потомки и родительское среднее | ½ VA = 1/2σ2A | b = VA /VP = σ2A / σ2P |
| Полусибсы | ¼ VA = 1/4σ2A | t = ¼ VA /VP = 1/4σ2A / σ2P |
| Полные сибсы | ½ VA + ¼ VD + VEc = = 1/2σ2 A + 1/4σ2 D + σ2 Ec | t = [ 1/2 VА + ¼ VD + VEc ]/ VP = = [1/2σ2 A + 1/4σ2 D + σ2 Ec ]/ σ2 P |
* Примечание: все обозначения даны ранее (см. табл.11.9). VEc = σ2 Ec – компонента средовой дисперсии, влияющая на сходство между полными сибсами.
Более подробно о вычислении доли ковариации между родственниками разного типа можно найти так же в упоминавшихся ранее работах Д. С. Фолконера.
Зная фенотипическую ковариацию родственников, можно рассчитать долю аддитивной генетической дисперсии, а коэффициенты регрессии или корреляции позволят определить довольно точно влияние аддитивной генетической дисперсии. Отметим, что доля общей дисперсии, представленная аддитивной генетической дисперсией представляет собой наследуемость. Более подробно об этой характеристике и ее использовании будет рассмотрено в соответствующей главе.
Связь между родственниками может выражаться или коэффициентом родства (коанцестри) или коэффициентом инбридинга. В настоящей главе рассматривается более общий случай, выражаемый коэффициентом родства. Инбридинг здесь будет затронут только частично, а более подробно рассмотрен в следующей главе.
Коанцестри
В любой отдельно взятой популяции древесных растений могут находиться особи, связанные различной степенью родства. Требуется какая-то мера, выражающая степень родства особей в популяции. Помимо коэффициента инбридинга, рассмотренного в предыдущем разделе, разными авторами для этого предлагается показатель, который называется коэффициент коанцестри, или коэффициент родства. Этот коэффициент отражает вероятность происхождения исследуемых индивидов от общего предка. В иностранной литературе он имеет разные названия: coefficient of coancestry, coefficient of consanguinity, coefficient of kinship (англ.), что означает коэффициент (кровного) родства, близости, сходства. Во французской литературе этот коэффициент родства обозначается как coefficient de parente '.
|
|
Определение этого показателя может быть представлено следующим образом. Коэффициент родства (коанцестри) - это вероятность того, что отдельные гены, взятые случайно от двух индивидуумов x и y, являются идентичными по происхождению (M. Lynch, B. Walsh, 1997; Dag Lindgren, 1999).
Из определения коанцестри видно, что в то время как коэффициент инбридинга (F) описывает родство между гомологичными генами индивидуума, коэффициент коанцестри (f) есть вероятность того, что ген, взятый случайно от одного индивидуума, будет идентичным по происхождению случайному гомологичному гену от другого индивидуума (T. J. Mullin, S. Bertrand, 1998).
Проблемами коанцестри для лесных древесных пород, а также других связанных с ними вопросов количественной генетики интенсивно занимается известный шведский генетик Даг Линдгрен и его школа (Dag Lindgren at al., 1997, a, b, c; Dag Lindgren, 1999 и др.). Эти подходы могут оказаться очень полезными как в программах селекции и семеноводства отдельных лесных древесных пород, так и лесовосстановления в целом.
Для дальнейшего изучения предмета необходимо дать или напомнить определения еще нескольких показателей.
Идентичный по происхождению (Identical by descends - IBD) означает, что гены одного и того же локуса являются копиями тех же самых первоначальных (оригинальных) генов какого-то предка, или анцестора (ancestor - предок по англ.).
Случай, когда оба гомологичных гена в той же самой зиготе являются идентичными по происхождению, называется инбридинг (F), или коэффициент инбридинга. Если две особи спариваются, их коанцестри становится инбридингом в их потомстве.
Основатель популяции является отправной точкой отсчета или вычислений. Если весь инбридинг и коанцестри основателя популяции известен, то инбридинг и коанцестри может быть рассчитан по родословной (педигри). Практично и удобно полагать инбридинг и коанцестри в "диком лесу" нулевыми и рассматривать основателей (плюсовые деревья) как образцы с дикого леса.
Самокoaнцестри - это коанцестри индивидуума с самим собой. Оно равно f = 0,5. Его можно представить, например, при допущении, что коанцестри в предыдущем поколении становится инбридингом в последующем при самоопылении.
Генный пул означает совокупность всех генов в популяции. Удобно рассматривать гены одного локуса. Пул генов не зависит от того, как популяция организована в зиготы.
Рассмотрим более подробно взаимоотношения между генами в популяции.
|
|
Популяция с N зиготами имеет 2 N генов в генном пуле
 |
Каждый ген имеет частоту 1/2 N
Вероятность выбрать тот же самый ген дважды равна 1/2 N.
Вероятность, что будут отобраны различные гены, равна (1-0,5/ N).
Гены могут быть идентичные по происхождению. Вероятность этого есть коанцестри.
 |
Генный пул структурирован по индивидуумам.
 |
Вероятность того, что гены в двух индивидуумах идентичны по происхождению, есть коанцестри между этими двумя индивидуумами.
Вероятность того, что различные гены одной и той же зиготы являются идентичными по происхождению, есть коэффициент инбридинга.
 |
Для самокоанцестри гены не нуждаются в различии. Если они различны - f=F, если те же самые - f=1, в среднем f=(1+F)/2.
В исследуемой популяции возможны три различных механизма определения генов, идентичных по происхождению:
1. Тот же самый ген отбирается дважды (дрейф);
2. Гены - есть гомологичные гены от того же самого индивидуума (инбридинг),
3. Гены происходят от различных индивидуумов (родство).
Оценки значений коанцестри спариваемых индивидуумов могут быть представлены в виде матрицы коанцестри (табл. 12.3).
Таблица 12.3
|
|
|

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

Археология об основании Рима: Новые раскопки проясняют и такой острый дискуссионный вопрос, как дата самого возникновения Рима...

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой...

Адаптации растений и животных к жизни в горах: Большое значение для жизни организмов в горах имеют степень расчленения, крутизна и экспозиционные различия склонов...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!