Аминокислоты, входящие в состав белков, являются a-аминокислотами. Все они принадлежат к L-ряду, а величина и знак оптического вращения зависят от природы радикалов аминокислот и значения рН раствора. В белках человека D-аминокислоты не обнаружены, однако они встречаются в клеточной стенке бактерий, в составе некоторых антибиотиков (актиномицинов).
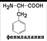
Аминокислоты отличаются друг от друга химической природой радикала R, который не участвует в образовании пептидной связи.
Современная рациональная классификация аминокислот основана на полярности радикалов:
Неполярные (гидрофобные)
Полярные (гидрофильные)
Ароматические (главным образом неполярные)
Отрицательно заряженные
Положительно заряженные
В некоторых белках обнаружены производные аминокислот. В белке соединительной ткани коллагене содержатся оксипролин и оксилизин. Дийодтирозин является основой структуры гормонов щитовидной железы.
Аминокислоты обладают общим свойством - амфотерностью (от греч amphoteros - двусторонний). В интервале рН 4,0-9,0 почти все аминокислоты существуют в форме биполяных ионов (цвиттерионов). Значение изоэлектрической точки аминокислоты (ИЭТ, рI) рассчитывается по формуле:
 .
.
Для моноаминодикарбоновых кислот рI рассчитывается как полусумма значений рK (таблица 1) a- и w-карбоксильных групп, для диаминомонокарбоновых кислот – как полусумма значений рK a- и w-аминогрупп.
Существуют заменимые аминокислоты (могут синтезироваться в организме человека), и незаменимые, которые в организме не образуются и должны поступать с пищей.
Незаменимые аминокислоты: валин, лейцин, изолейцин, лизин, метионин, треонин, триптофан, фенилаланин.
Заменимые аминокислоты: глицин, аланин, аспарагин, аспартат, глутамин, глутамат, пролин, серин.
Условно заменимые (могут синтезироваться в организме из других аминокислот): аргинин (из цитруллина), тирозин (из фенилаланина), цистеин (из серина), гистидин (при участии глутамина).
Содержание различных аминокислот в белках неодинаково.
Для открытия в биологических объектах и количественного определения аминокислот используют реакцию с нингидрином.
3.Специфичность первичной структуры белка. Особенности образования пептидной связи. Определяющая роль первичной структуры в формировании более высоких уровней организации белковой молекулы.
Первичная структура белка – последовательность расположения аминокислотных остатков в полипептидной цепи. В белках отдельные аминокислоты связаны друг с другом пептидными связями, возникающими при взаимодействии a-карбоксильных и a-аминогрупп аминокислот.
Полипептидная цепь состоит из регулярно повторяющихся участков, образующих остов молекулы, и вариабельных участков – боковых радикалов аминокислотных остатков. Началом полипептидной цепи считают конец, несущий свободную аминогруппу (N-конец), а заканчивается полипептидная цепь свободной карбоксильной группой (С-конец). Как правило, при изображении формулы пептида N-конец располагают слева, а С-конец — справа
Первичная структура каждого индивидуального белка закодирована в молекуле ДНК (в участке, называемом геном) и реализуется в ходе транскрипции (переписывания информации на мРНК) и трансляции (синтез первичной структуры белка). Следовательно, первичная структура белков индивидуального человека - наследственно передаваемая от родителей детям информация, определяющая особенности строения белков данного организма, от которых зависит функция имеющихся белков
Каждый из примерно 100 000 индивидуальных белков в организме человека имеет уникальную первичную структуру. В молекулах одного типа белка (например, альбумина) одинаковое чередование аминокислотных остатков, что отличает альбумин от любого другого индивидуального белка.
Последовательность аминокислотных остатков в пептидной цепи можно рассматривать как форму записи информации. Эта информация определяет пространственную укладку линейной пептидной цепи в более компактную трехмерную структуру, называемую конформацией белка. Процесс формирования функционально активной конформации белка носит название фолдинг.
Конформация белков. Свободное вращение в пептидном остове возможно между атомом азота пептидной группы и соседним α-углеродным атомом, а также между α-углеродным атомом и углеродом карбонильной группы. Вследствие взаимодействия функциональных групп аминокислотных остатков первичная структура белков может приобретать более сложные пространственные структуры. В глобулярных белках различают два основных уровня укладки конформации пептидных цепей: вторичную и третичную структуры.
4.Вторичная структура белка. Связи, стабилизирующие вторичную структуру, α-спираль. Факторы, нарушающие спирализацию. β-складчатая структура, особенности конформационного строения.
Вторичная структура представляет собой способ укладки полипептидной цепи в упорядоченную структуру благодаря образованию водородных связей между пептидными группами одной цепи или смежными полипептидными цепями. По конфигурации вторичные структуры делятся на спиральные (α-спираль) и слоисто-складчатые (β-структура и кросс-β-форма).
α-Спираль. Это разновидность вторичной структуры белка, имеющая вид регулярной спирали, образующейся благодаря межпептидным водородным связям в пределах одной полипептидной цепи. Модель строения α-спирали (рис. 2), учитывающая все свойства пептидной связи, была предложена Полингом и Кори. Основные особенности α-спирали:
· спиральная конфигурация полипептидной цепи, имеющая винтовую симметрию;
· образование водородных связей между пептидными группами каждого первого и четвертого аминокислотных остатков;
· регулярность витков спирали;
· равнозначность всех аминокислотных остатков в α-спирали независимо от строения их боковых радикалов;
· боковые радикалы аминокислот не участвуют в образовании α-спирали.
Внешне α-спираль похожа на слегка растянутую спираль электрической плитки. Регулярность водородных связей между первой и четвертой пептидными группами определяет и регулярность витков полипептидной цепи. Высота одного витка, или шаг α-спирали, равна 0,54 нм; в него входит 3,6 аминокислотных остатка, т. е. каждый аминокислотный остаток перемещается вдоль оси (высота одного аминокислотного остатка) на 0,15 нм (0,54:3,6 = 0,15 нм), что и позволяет говорить о равнозначности всех аминокислотных остатков в α-спирали. Период регулярности α-спирали равен 5 виткам или 18 аминокислотным остаткам; длина одного периода составляет 2,7 нм. Рис. 3. Модель а-спирали Полинга—Кори
β-Структура. Это разновидность вторичной структуры, которая имеет слабо изогнутую конфигурацию полипептидной цепи и формируется с помощью межпептидных водородных связей в пределах отдельных участков одной полипептидной цепи или смежных полипептидных цепей. Ее называют также слоисто-складчатой структурой. Имеются разновидности β-структур. Ограниченные слоистые участки, образуемые одной полипептидной цепью белка, называют кросс-β-формой (короткая β-структура). Водородные связи в кросс-β-форме образуются между пептидными группами петель полипептидной цепи. Другой тип — полная β-структура — характерен для всей полипептидной цепочки, которая имеет вытянутую форму и удерживается межпептидными водородными связями между смежными параллельными полипептидными цепями (рис. 3). Эта структура напоминает меха аккордеона. Причем возможны варианты β-структур: они могут быть образованы параллельными цепями (N-концы полипептидных цепей направлены в одну и ту же сторону) и антипараллельными (N-концы направлены в разные стороны). Боковые радикалы одного слоя помещаются между боковыми радикалами другого слоя.
В белках возможны переходы от α-структур к β-структурам и обратно вследствие перестройки водородных связей. Вместо регулярных межпептидных водородных связей вдоль цепи (благодаря им полипептидная цепь скручивается в спираль) происходит раскручивание спирализованных участков и замыкание водородных связей между вытянутыми фрагментами полипептидных цепей. Такой переход обнаружен в кератине — белке волос. При мытье волос щелочными моющими средствами легко разрушается спиральная структура β-кератина и он переходит в α-кератин (вьющиеся волосы распрямляются).
Разрушение регулярных вторичных структур белков (α-спирали и β-структур) по аналогии с плавлением кристалла называют "плавлением" полипептидов. При этом водородные связи рвутся, и полипептидные цепи принимают форму беспорядочного клубка. Следовательно, стабильность вторичных структур определяется межпептидными водородными связями. Остальные типы связей почти не принимают в этом участия, за исключением дисульфидных связей вдоль полипептидной цепи в местах расположения остатков цистеина. Короткие пептиды благодаря дисульфидным связям замыкаются в циклы. Во многих белках одновременно имеются α-спиральные участки и β-структуры. Природных белков, состоящих на 100% из α-спирали, почти не бывает (исключение составляет парамиозин — мышечный белок, на 96-100% представляющий собой α-спираль), тогда как у синтетических полипептидов 100%-ная спирализация.
Другие белки имеют неодинаковую степень спирализации. Высокая частота α-спиральных структур наблюдается у парамиозина, миоглобина, гемоглобина. Напротив, у трипсина, рибонуклеазы значительная часть полипептидной цепи укладывается в слоистые β-структуры. Белки опорных тканей: кератин (белок волос, шерсти), коллаген (белок сухожилий, кожи), фиброин (белок натурального шелка) имеют β-конфигурацию полипептидных цепей. Разная степень спирализации полипептидных цепей белков говорит о том, что, очевидно, имеются силы, частично нарушающие спирализацию или "ломающие" регулярную укладку полипептидной цепи. Причиной этого является более компактная укладка полипептидной цепи белка в определенном объеме, т. е. в третичную структуру.
5.Третичная структура белка. Связи, стабилизирующие третичную структуру (ковалентные, ионные, гидрофобные, водородные, Ван-дер-Ваальса).
Третичной структурой белка называется способ укладки полипептидной цепи в пространстве. По форме третичной структуры белки делятся в основном на глобулярные и фибриллярные. Глобулярные белки чаще всего имеют эллипсовидную форму, а фибриллярные (нитевидные) белки — вытянутую (форма палочки, веретена).
Однако конфигурация третичной структуры белков еще не дает основания думать, что фибриллярные белки имеют только β-структуру, а глобулярные α-спиральные. Есть фибриллярные белки, имеющие спиральную, а не слоисто-складчатую вторичную структуру. Например, α-кератин и парамиозин (белок запирательной мышцы моллюсков), тропомиозины (белки скелетных мышц) относятся к фибриллярным белкам (имеют палочковидную форму), а вторичная структура у них — α-спираль; напротив, в глобулярных белках может быть большое количество β-структур.
Спирализация линейной полипептидной цепи уменьшает ее размеры примерно в 4 раза; а укладка в третичную структуру делает ее в десятки раз более компактной, чем исходная цепь.
Связи, стабилизирующие третичную структуру белка. В стабилизации третичной структуры играют роль связи между боковыми радикалами аминокислот. Эти связи можно разделить на:
· сильные (ковалентные) К полярным связям относятся водородные и ионные. Водородные связи, как обычно, возникают между группой —NН2, — ОН или —SН бокового радикала одной аминокислоты и карбоксильной группой другой. Ионные, или электростатические, связи образуются при контакте заряженных групп боковых радикалов —NН+3 (лизина, аргинина, гистидина) и —СОО- (аспарагиновой и глутаминовой кислот). Неполярные, или ван-дер-ваальсовы, связи образуются между углеводородными радикалами аминокислот. Гидрофобные радикалы аминокислот аланина, валина, изолейцина, метионина, фенилаланина в водной среде взаимодействуют друг с другом. Слабые ван-дер-ваальсовы связи способствуют формированию гидрофобного ядра из неполярных радикалов внутри белковой глобулы. Чем больше неполярных аминокислот, тем большую роль в укладке полипептидной цепи играют ван-дер-ваальсовы связи.
· слабые (полярные и ван-дер-ваальсовы) К полярным связям относятся водородные и ионные. Водородные связи, как обычно, возникают между группой —NН2, — ОН или —SН бокового радикала одной аминокислоты и карбоксильной группой другой. Ионные, или электростатические, связи образуются при контакте заряженных групп боковых радикалов —NН+3 (лизина, аргинина, гистидина) и —СОО- (аспарагиновой и глутаминовой кислот). Неполярные, или ван-дер-ваальсовы, связи образуются между углеводородными радикалами аминокислот. Гидрофобные радикалы аминокислот аланина, валина, изолейцина, метионина, фенилаланина в водной среде взаимодействуют друг с другом. Слабые ван-дер-ваальсовы связи способствуют формированию гидрофобного ядра из неполярных радикалов внутри белковой глобулы. Чем больше неполярных аминокислот, тем большую роль в укладке полипептидной цепи играют ван-дер-ваальсовы связи.
Многочисленные связи между боковыми радикалами аминокислот определяют пространственную конфигурацию белковой молекулы.
Особенности организации третичной структуры белка. Конформация третичной структуры полипептидной цепи определяется свойствами боковых радикалов входящих в нее аминокислот (которые не оказывают заметного влияния на формирование первичной и вторичной структур) и микроокружением, т. е. средой. При укладке полипептидная цепь белка стремится принять энергетически выгодную форму, характеризующуюся минимумом свободной энергии. Поэтому неполярные R-группы, "избегая" воды, образуют как бы внутреннюю часть третичной структуры белка, где расположена основная часть гидрофобных остатков полипептидной цепи. В центре белковой глобулы почти нет молекул воды. Полярные (гидрофильные) R-группы аминокислоты располагаются снаружи этого гидрофобного ядра и окружены молекулами воды. Полипептидная цепь причудливо изгибается в трехмерном пространстве. При ее изгибах нарушается вторичная спиральная конформация. "Ломается" цепь в слабых точках, где находятся пролин или гидроксипролин, поскольку эти аминокислоты более подвижны в цепи, образуя только одну водородную связь с другими пептидными группами. Другим местом изгиба является глицин, R-группа которого мала (водород). Поэтому R-группы других аминокислот при укладке стремятся занять свободное пространство в месте нахождения глицина. Ряд аминокислот - аланин, лейцин, глутамат, гистидин - способствуют сохранению устойчивых спиральных структур в белке, а такие, как метионин, валин, изолейцин, аспарагиновая кислота, благоприятствуют образованию β-структур. В молекуле белка с третичной конфигурацией встречаются участки в виде α-спиралей (спирализованные), β-структур (слоистые) и беспорядочного клубка. Только правильная пространственная укладка белка делает его активным; нарушение ее приводит к изменению свойств белка и потере биологической активности.