Первичный Посредник
↓ ↓ ↓
Аденилатциклаза Гуанилатциклаза Фосфолипаза С

 ↓ ↓ ↓
↓ ↓ ↓
АТФ ГТФ Фосфатидолинозитол-4,5- дифосфат
↓ ↓ ↓ ↓
цАМФ цГМФ Диацилглицерол Инозитолтрифосфат
↓ ↓ ↓ ↓
Са++ Са++ Са++ Са++
Эффекты:
 1) фосфолирирование белков, конформация белков и ответ клетки;
1) фосфолирирование белков, конформация белков и ответ клетки;
2) связь ионов Са с белками.
Рецепторы. По локализации различают мембранные, цитоплазматические и ядерные рецепторы. По другой классификации все рецепторы можно разделить на быстроотвечающие (в пределах миллисекунд) и медленноотвечающие, в пределах нескольких минут или даже часов, что характерно для гормонов, передающих сигнал на внутриклеточные рецепторы. Рецепторы первого типа - интегральные олигомерные белки, содержащие субъединицу, имеющую центр для связывания сигнальной молекулы и центральный ионный канал.
Рецепторы второго типа, локализованные в мембранах и не связанные с каналами, подразделяют на 2 большие группы: каталитические рецепторы, обладающие собственной тирозинкиназной или гуанилатциклазной активностью, и рецепторы, взаимодействующие через G-белок с мембранным ферментом. Связывание лиганда (например, гормона) с рецептором на наружной стороне клеточной мембраны приводит к изменению активности цитоплазматического фермента, который, в свою очередь, инициирует клеточный ответ, т.е. через мембрану переносится информация, а не заряды или растворённые молекулы.
В случае цитоплазматических рецепторов через мембрану проходит гормон, а информация о присутствии гормона в клетке с помощью рецептора передаётся в ядро.
Различные клетки организма в зависимости от выполняемых ими функций имеют определённый набор рецепторов. В мембране одной клетки может быть более десятка разных типов рецепторов. Взаимодействуя с рецептором, внеклеточные химические посредники оказывают влияние на метаболизм и функциональное состояние (пролиферация, секреция и т.д.) клеток-мишеней.
Взаимодействие гормонов с рецепторами, сопряженными с G-белками, приводит к активации аденилатциклазной или инозитолфосфатной регуляторных систем
50. Трансмембранная передача сигналов на примере аденилатциклазной мессенджерной системы.
Наиболее изученной является аденилатциклазная мессенджерная система (рис. 29).В нее входят рецептор гормона, G-белок, фермент аденилатциклаза, цАМФ-зависимая протеинкиназа, фосфодиэстераза.
Рецепторы гидрофильных гормонов, таких как адреналин, расположены на мембране. Связывание гормона (первичного мессенджера) с рецептором приводит к структурным изменениям внутриклеточного домена рецептора, что обеспечивает взаимодействие рецептора с ГТФ-связывающим белком (G-белком). G-белок представляет собой смесь 2 типов белков: активного Gs (от англ. stimulatory) и ингибиторного Gi. В составе каждого из них имеется три субъединицы (α, β и γ). Функция G-белка - проведение гормонального сигнала на уровне плазматической мембраны. В состоянии покоя G-белок связан с ГДФ. Гормонрецепторный комплекс переводит Gs-белок в активированное состояние, ГДФ замещается на ГТФ, что приводит к отделению субъединицы α, которая активирует аденилатциклазу. В отсутствие G-белка аденилатциклаза практически неактивна.
Аденилатциклаза- интегральный белок плазматических мембран, его активный центр ориентирован в сторону цитоплазмы. Аденилатциклаза катализирует реакцию синтеза из АТФ цАМФ (вторичного мессенджера).
Протеинкиназа А катализирует фосфорилирование внутриклеточных ферментов или белков-мишеней, изменяя их активность. Неактивная протеинкиназа А состоит из 4 субъединиц. Под действием цАМФ она переходит в активную форму за счет диссоциации субъединиц и фосфорилирует белок за счет АТФ. Процесс фосфорилирования-дефосфорилирования белков при участии протеинкиназ является общим фундаментальным механизмом действия вторичных мессенджеров внутри клетки.
Фосфодиэстераза вызывает распад цАМФ и тем самым прекращает действие сигнала.
51.  Трансмембранная передача сигналов на примере инозитолфосфатной мессенджерной системы.
Трансмембранная передача сигналов на примере инозитолфосфатной мессенджерной системы.
Инозитолфосфатная система включает три основных мем-бранных белка: рецептор гормона, фосфолипазу С и белок Gplc, активирующий фосфолипазу.
Связывание гормона с рецептором приводит к изменению его конфор-мации и увеличению сродства с белком Gplc. Он представляет собой тример, состоящий из субъединиц α, β и γ. При участии ГТФ комплекс «гормон - рецептор – G-белок» диссоциирует с освобождением -субъединицы. Она взаимодействует с фосфолипазой С и активирует ее. Субстрат этого фермента – фосфатидилинозитол-4,5-бисфосфат (ФИФ). В инозитолфосфатной системе в роли вторичных мессенджеров участвуют инозитол-1,4,5-трисфосфат (ИФ-3), диацилглицерин (ДАГ) и ионы Са2+. ИФ-3 выходит в цитозоль. ДАГ остается в мембране и участвует в активации фермента протеинкиназы С. ИФ-3 связывается с кальциевым каналом мембраны ЭПР, канал открывается, и ионы Са2+ поступают в цитозоль. Увеличивается скорость взаимодействия ионов Са2+ с неактивной протеинкиназой С и белком кальмодулином.
На внутренней стороне мебраны образуется комплекс «протеинкиназа С - Са2+ - ДАГ – фосфолипаза С». Происходит активирование протеинкиназы С, которая фосфорилирует ферменты по остаткам серина и треонина, меняя их активность.
Кальмодулин имеет 4 центра для связывания Са2+. Взаимодействие комплекса «кальмодулин - Са2+» с ферментами приводит к их активации.
Аденилатциклазная и инозитолфосфатная системы регулируют большое количество разных клеточных процессов. Эффект этих систем проявляется очень быстро.
Стероидные и тиреоидные гормоны обладают липофильнымисвой-ствами и легко проходят через клеточные мембраны. Их рецепторы находятся в цитозоле или в ядре клетки (внутриклеточные рецепторы).
Если рецептор гормона находится в цитозоле, там же происходит образование комплекса гормона с рецептором, который затем поступает в ядро. Рецепторы ряда гормонов расположены в ядре клетки, тогда комплекс «гормон – рецептор» может образоваться непосредственно в ядре.
В ядре комплекс гормон-рецептор взаимодействует с регуляторной нуклеотидной последовательностью ДНК, что приводит к изменению скорости транскрипции структурных генов и, следовательно, скорости трансляции. В результате изменяется количество белков, которые участвуют в метаболизме и влияют на функциональное состояние клетки.
Эффекты гормонов, которые передают сигнал через внутриклеточные рецепторы, нельзя наблюдать сразу, так как на протекание матричных процессов (транскрипцию и трансляцию) требуются часы.
Гормоны обеспечивают коммуникацию (обмен информацией) между разными клетками и органами. В результате действия этих механизмов до-стигается координация метаболизма и функций разных клеток и органов и адекватная реакция организма на изменения внешней среды.
В роли внеклеточных сигналов могут действовать не только гормоны, но и ряд других веществ - цитокины, биогенные амины, нейромедиаторы и др.
52. Передача гормональных сигналов на примере стероидных гормонов.
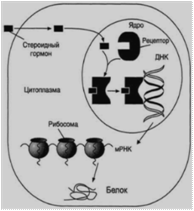 Липофильные гормоны, за исключением ЙТ и ретиноата, являются производными холестерина. После секреции свободный гормон диффундирует сквозь плазматическую мембрану и связывается с рецептором либо в гиалоплазме, либо в ядре. Возникший гормон-рецепторный комплекс связывается со специфическим участком ДНК, который называется гормон-реактивным элементом (ГРЭ). Он функционально аналогичен энхансеру или сайленсеру, и в результате меняется экспрессия гена (то есть активируются или ингибируются специфические гены). Например, происходит увеличение матки во время беременности, рост молочной железы перед и во время лактации.Между двумя группами липофильных гормонов есть существенные различия в локализации их рецепторов (рис.11). Рецепторы стероидных гормонов находятся в цитозоле в неактивном комплексе с шаперонами, после их диссоциации гормон-рецепторный комплекс перемещается в ядро и там взаимодействует с ГРЭ. Рецепторы йодтиронинов и активных форм витаминов D (кальцитриол) и А (ретиноат) постоянно находятся в ядре клетки. Свободному гормону приходится проникать не только в клетку, но и в ядро и взаимодействовать с соответствующим ГРЭ ДНК.
Липофильные гормоны, за исключением ЙТ и ретиноата, являются производными холестерина. После секреции свободный гормон диффундирует сквозь плазматическую мембрану и связывается с рецептором либо в гиалоплазме, либо в ядре. Возникший гормон-рецепторный комплекс связывается со специфическим участком ДНК, который называется гормон-реактивным элементом (ГРЭ). Он функционально аналогичен энхансеру или сайленсеру, и в результате меняется экспрессия гена (то есть активируются или ингибируются специфические гены). Например, происходит увеличение матки во время беременности, рост молочной железы перед и во время лактации.Между двумя группами липофильных гормонов есть существенные различия в локализации их рецепторов (рис.11). Рецепторы стероидных гормонов находятся в цитозоле в неактивном комплексе с шаперонами, после их диссоциации гормон-рецепторный комплекс перемещается в ядро и там взаимодействует с ГРЭ. Рецепторы йодтиронинов и активных форм витаминов D (кальцитриол) и А (ретиноат) постоянно находятся в ядре клетки. Свободному гормону приходится проникать не только в клетку, но и в ядро и взаимодействовать с соответствующим ГРЭ ДНК.
53. Сущность понятий: метаболизм, анаболизм, катаболизм. Три фазы катаболизма (переваривание, специфические и общие пути катаболизма), их назначение, энергетическая ценность. Понятие о ключевых метаболитах организма человека (ацетил-КоА, ПВК).
Метаболизм (от греч. «превращение, изменение») или обмен веществ - совокупность химических реакций, протекающих в живом организме, и обеспечивающих его жизнедеятельность.
Катаболизм – совокупность процессов, связанных с распадом веществ, их окислением и выведением из организма продуктов распада (СО2, Н2О, мочевина), при этом выделяется энергия (экзергонические реакции).
Органоиды катаболической системы – митохондрии, лизосомы, пероксисомы.
Анаболизм – совокупность реакций синтеза необходимых для оранизма веществ, их использование для роста, развития и жизнедеятельности организма; при этом, как правило, затрачивается энергия (эндергонические реакции).
Органоиды анаболической системы – эндоплазматический ретикулум, комплекс Гольджи, рибосомы.
I этап катаболизма – гидролитический. Под воздействием гидролаз в пищеварительном тракте белки, жиры, углеводы распадаются на соответствующие мономеры (рис. 32).
II этап – специфические пути катаболизма. Мономеры основных пищевых веществ при участии ферментов, специфичных для каждого метаболита, превращаются в основном в два метаболита - пировиноградную кислоту (ПВК) и ацетил-KоА. Ацетил-KоА (ацетилкоэнзим А) - макроэргический продукт конденсации коэнзима А с уксусной кислотой.
Реакции специфических путей катаболизма протекают внутри клеток. На этом этапе высвобождается 1/3 энергии питательных веществ.
III этап - общий путь катаболизма. После образования ПВК дальнейший путь распада веществ до СО2 и Н2О происходит одинаково в общем пути катаболизма. Он включает два процесса:
1) окислительное декарбоксилирование ПВК;
2) цикл Кребса (цикл трикарбоновых кислот (ЦТК), цитратный цикл).
В общем пути катаболизма образуются первичные доноры водорода для цепи переноса электронов (дыхательной цепи). Реакции общего пути катаболизма происходят в матриксе митохондрий, и восстановленные коферменты передают водород непосредственно на компоненты дыхательной цепи, расположенные во внутренней мембране митохондрий, где образуется АТФ.
На этом этапе высвобождается 2/3 энергии питательных веществ.
Метаболиты - продукты метаболизма каких-либо соединений.
Ацетил-кофермент А, ацетил-коэнзим А, сокращённо ацетил-КоА — важное соединение в обмене веществ, используемое во многих биохимических реакциях. Его главная функция — доставлять атомы углерода с ацетил-группой в цикл трикарбоновых кислот, чтобы те были окислены с выделением энергии. По своей химической структуре ацетил-КоА — тиоэфир между коферментом А (тиолом) и уксусной кислотой (носителем ацильной группы). Ацетил-КоА образуется во время второго шага кислородного клеточного дыхания, декарбоксилирования пирувата, который происходит в матриксе митохондрии. Ацетил-КоА затем поступает в цикл трикарбоновых кислот. Ацетил-КоА — важный компонент биологического синтеза нейротрансмиттера ацетилхолина. Холин, в соединении с ацетил-КоА, катализируется ферментом холинацетилтрансферазой, чтобы образовать ацетилхолин и коэнзим А. 
ПВК является конечным продуктом метаболизма глюкозы в процессе гликолиза. Одна молекула глюкозы превращается при этом в две молекулы пировиноградной кислоты. Дальнейший метаболизм пировиноградной кислоты возможен двумя путями — аэробным и анаэробным.
В условиях достаточного поступления кислорода пировиноградная кислота превращается в ацетил-кофермент А, являющийся основным субстратом для серии реакций, известных как цикл Кребса, или дыхательный цикл, цикл трикарбоновых кислот. Пируват также может быть превращён в анаплеротической реакции в оксалоацетат. Оксалоацетат затем окисляется до углекислого газа и воды.
Если кислорода недостаточно, пировиноградная кислота подвергается анаэробному расщеплению с образованием молочной кислоты у животных и этанола у растений. При анаэробном дыхании в клетках пируват, полученный при гликолизе, преобразуется в лактат при помощи фермента лактатдегидрогеназы и NADP в процессе лактатной ферментации, либо в ацетальдегид и затем в этанол в процессе алкогольной ферментации.. Пируват может быть превращён обратно в глюкозу в процессе глюконеогенеза, или в жирные кислоты или энергию через ацетил-КоА, в аминокислоту аланин, или в этанол. Таким образом пируват объединяет несколько ключевых метаболических процессов клетки и является универсальной «метаболической валютой» живой клетки.
Общая схема окислительного декарбоксилирования пировиноградной кислоты (ПВК), локализация процесса. Строение пируватдегидрогеназного комплекса. Регуляция процесса окислительного декарбоксилирования ПВК.
Окислительное декарбоксилирование пировиноградной кислоты - многостадийный процесс, который катализирует пируватдегидрогеназный комплекс - митохондриальный мультиферментный комплекс, соединенный с внутренней мембраной со стороны матрикса. ПВК поступает к комплексу из матрикса, и туда же освобождаются продукты реакции.
Пируватдегидрогеназный комплекс включает три фермента (пируватдегидрогеназа (Е1), ацетилтрансфераза (Е2), дегидрогеназа дигидролипоевой кислоты (Е3)) и пять коферментов (НАД+, ФАД, тиаминпирофосфат, липоевая кислота, кофермент А (рис. 36)).
Тиаминпирофосфат связан с пируватдегидрогеназой (Е1), липоевая кислота - с ацетилтрансферазой (Е2), ФАД - с дегидрогеназой дигидролипоевой кислоты (Е3). Кофермент А и НАД+ находятся в свободно растворенном состоянии.
В состав пируватдегидрогеназного комплекса входит примерно по три десятка молекул Е1 и Е2 и 10 молекул Е3. Комплекс работает как конвейер: промежуточные продукты не освобождаются в раствор, а передаются от фермента к ферменту.
Первую реакцию катализирует Е1, субстратами являются ПВК и дегидролипоевая кислота, являющаяся простетической группой Е2. От ПВК отщепляется карбоксильная группа и образуется СО2, а ацетильный остаток соединяется с атомом серы липоевой кислоты в составе ацетилтрансферазы. Образуется ацетиллипоат-Е2.
Во второй реакции ацетилтрансфераза (Е2) катализирует перенос ацетильного остатка, соединенного с его простетической группой, на коэнзим А. Продукты этой реакции - дигидролипоевая кислота в составе Е2 и ацетил-KоА.
В третьей реакции происходит дегидрирование дигидролипоевой кислоты в составе ацетилтрансферазы при воздействии фермента Е3 (дегидрогеназа дигидролипоевой кислоты), содержащего ФАД. ФАД передает водород на НАД+. Образуются НАДН+Н+ и дегидролипоевая кислота в составе Е2. Последний фермент снова вступает в окислительное декарбоксилирование ПВК.
Ацетил-KоА (продукт второй реакции) затем окисляется в цикле Кребса. Водород с НАДН (продукт третьей реакции) поступает в дыхательную цепь, где образуется АТФ.
Энергетический выход окислительного декарбоксилирования пирувата – 3 АТФ.
Суммарное уравнение процесса:
СH3-CО-COOH + HS-KoA + НАД+ ® СH3-CО-S-KoA + НАДН + Н+ + СО2.
По механизму «обратной связи» работу пируватдегидрогеназного комплекса ингибируют конечные продукты окислительного декарбоксилирования - ацетил-KоА, НАДН+Н+, а также АТФ. Увеличивают активность комплекса пировиноградная кислота и АДФ.
Также имеет место гормональная регуляция: инсулин увеличивает активность пируатдегидрогеназного комплекса, глюкагон - снижает.








