Томас Морган в 1910-1911гг., проводя опыты на дрозофиле (Drosofila melanogaster), установил ряд общих генетических закономерностей. Он работал с уксусной мушкой (Drosofila melanogaster), имеющей 4 пары хромосом, при этом 3 пары аутосом идентичны у самца и самки, а по одной паре хромосом они различаются. Это, так называемые, половые хромосомы. У самки имеется 6 аутосом + Х Х – половые хромосомы. У самца, кроме 6 аутосом, имеются ещё две половые хромосомы Х и Y. Самец производит гаметы двух типов: половина гамет имеют 3 аутосомы + Х -хромосому; другая половина гамет имеют 3 аутосомы + Y -хромосому. Гаметы самки в отношении половых хромосом – одинаковые, в результате мейотического деления каждая яйцеклетка получает одну Х -хромосому из двух.
Если гены находятся в половых хромосомах, то наследование признаков, определяемое этими генами, будет зависеть от поведения половых хромосом в мейозе. Считается, что Y -хромосома, в отличие от Х -хромосомы наследственно инертна и не контролирует большинства признаков. Поэтому гены, находящиеся в Х -хромосоме, не имеют аллелей в Y -хромосоме. Если Х и Y -хромосомы находятся рядом, то рецессивные гены в Х -хромосоме, проявляются так, как обычно бывает у рецессивных гомозигот.
Т. Морган скрестил муху «дикого» типа, имеющую красные глаза RR c мутантным белоглазым самцом rr. В F 1 у всех мух были красные глаза и расщепление по полу 1:1.В F 2 расщепление по полу сохранялось 1:1, при этом все самки были красноглазыми, а у самцов происходило расщепление по этому признаку в равном соотношении на красноглазых и белоглазых. (Рис. 5.3.)
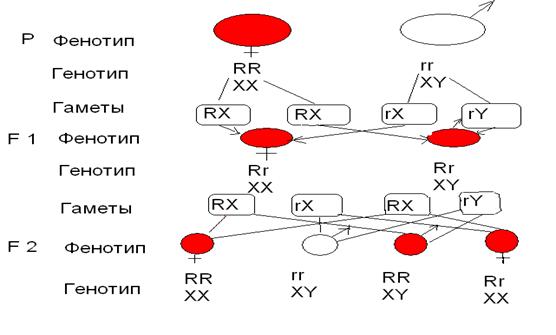
Рис. 5.3. Наследование признаков, сцепленных с полом, у Drosophila melanogaster при скрещивании красноглазой самки и белоглазого самца.
В реципрокном скрещивании, когда самка была белоглазой, а самец кросноглазый в F 1 наблюдалось расщепление по цвету глаз: самки были красноглазыми, как отец, а самцы – белоглазыми, как мать. В F 2 среди самок и самцов половина особей имела красные глаза, половина – белые. (Рис. 5.4.)
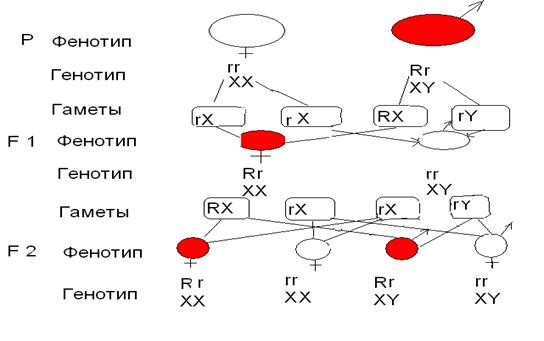
Рис. 5.4. Наследование признаков, сцепленных с полом, у Drosophila melanogaster при скрещивании белоглазой самки и красноглазого самца (крисс-кросс)
Морган показал, что все эти результаты можно объяснить, если предположить, что, во-первых, ген, определяющий цвет глаз, расположен в половой хромосоме (Х -хромосоме) и, во-вторых, что половая хромосома самцов (Y -хромосома) не содержит этот ген. Самки получают по одной Х -хромосоме от отца и от матери и передают их как дочерям, так и сыновьям. Самцы же получают Х -хромосому от матери и передают её лишь дочерям (рис. 5.3 и 5.4). В соответствии с этим признаки, определяемые генами, расположенными в Х -хромосоме, наследуются «крест-накрест»: самцы передают эти признаки внукам лишь через дочерей, но не через сыновей.
Морган пришел к выводу, что белый цвет глаз у дрозофилы – это признак, сцепленный с полом. То есть, ген, определяющий этот признак, расположен в Х -хромосоме. Самцы называются гемизиготными по генам, расположенным в Х -хромосоме, поскольку в отношении этих генов они не являются ни гомо, ни гетерозиготными.
Наследование признаков, сцепленных с полом, полностью соответствует распределению половых хромосом в мейозе и сочетанию их при оплодотворении. На основании этого был сделан вывод, что гены, определяющие данные признаки, действительно находятся в половых хромосомах.
Тип скрещивания, когда гены матери наследует сын, а гены отца — дочь, называется крисс-кросс (крест-накрест), а особи, гетерогометные по половым хромосомам - гемизиготными. У человека изучено около 150 признаков, сцепленных с полом. В Х – хромосоме локализовано около 300 генов, вызывающих наследственные заболевания.
(Е.К. Гинтер, 2003; Н.П. Бочков, 2006). Наиболее известными из них являются дальтонизм, гемофилия, миопатия Дюшена, Х- сцепленный ихтиоз, синдром ломкой хромосомы Х с умственной отсталостью, гидроцефалии, пигментная дистрофия сетчатки и многие другие заболевания. Классическим примером наследования сцепленного с полом является наследование полулетального Х – сцепленного гена гемофилии в королевских семьях Европы, которые восходят к английской королеве Виктории. Она была гетерозиготной по гену гемофилии и передала его через дочерей внукам и правнукам целого ряда царственных семей. Этот ген вызывает несвёртываемость крови, и даже небольшая ранка может привести к летальному исходу. Заболевание проявляется у мальчиков. От гемофилии погибли три испанских инфанта (Альфонсо, Гонзалес, Жуан), матерью которых была внучка королевы Виктории – Виктория – Евгения Баттенбергская. Её родные братья Леопольд и Морис Баттенбергские страдали гемофилией, в то время как их отец Генрих Баттенбергский был здоров.
В прусскую монархическую ветвь ген гемофилии внесла дочь королевы Виктории – Алиса через свою дочь – Ирену. Сестра Ирены Алиса (Александра Фёдоровна) передала ген гемофилии своему сыну – цесаревичу Алексею, наследнику российского престола. И Ирена и Александра Фёдоровна осознавали свою причастность к заболеваниям сыновей, так как их брат Фридрих Вильгельм умер от подобного же заболевания. Анализ генеалогического древа царствующих семей Европы показывает, что сыновья, больные гемофилией, появляются в браках, где оба родителя по данному признаку нормальны. Передатчиком болезни в таких случаях является мать, гетерозиготная по рецессивному гену гемофилии. Если мужчины, больные гемофилией, достигнут половозрелости и вступят в брак, то все их дочери окажутся геторозиготными по гену гемофилии, а затем передадут этот ген половине своих сыновей. В гомозиготном состоянии ген гемофилии для женщин летален. Явление гемофилии известно у животных, например, у собак. (А.В. Бакай и др.,2006)
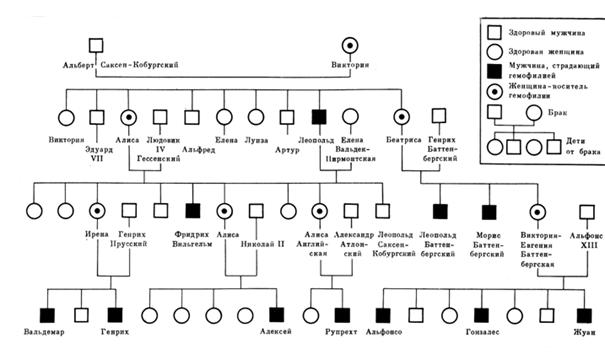
Рис.5.5. Генеалогическое древо царствовавших семей Европы, иллюстрирующее наследование гена гемофилии, локализованного в Х- хромосоме человека. (Н.Н.Дубинин, 1976).
При заболевании мышечной дистрофией (миопатия Дюшена) мужчины не имеют потомства, так как она проявляется в раннем детстве и ведёт к инвалидности и ранней смерти. Женщины, гетерозиготные по гену этого заболевания, здоровы.
Известны нарушения цветового зрения связанные с Х – сцепленным наследованием, называется это нарушение дальтонизмом по имени французского физика Дальтона, имеющего дефект с восприятием красно-зеленого цвета.
В отличие от первых двух заболеваний (гемофилии и мышечной дистрофии), где женщины являются носителями признака, ген дальтонизма не является летальным и в гомозиготном состоянии проявляться у женщин. Если женщина, страдающая цветовой слепотой к красному цвету, выходит замуж за мужчину с нормальным зрением, то у их дочерей зрение нормальное, а сыновья страдают цветовой слепотой. Если дальтоник женится на женщине с нормальным восприятием цвета, то все дети от этого брака не будут дальтониками. Возможны случаи, когда у родителей с нормальным зрением, половина сыновей оказываются поражёнными цветовой слепотой к красному цвету.
Распространение цветовой слепоты к красному цвету среди населения различно для мужчин и женщин. В США около 8% мужчин имеют такую патологию, в то время как женщины только 0,5%. В настоящее время доказано, что женщины-носительницы мутации красно-зелёной цветовой слепоты имеют мозаичную сетчатку с нормальными и мутантными клетками и сохраняют способность воспринимать цвета. Впервые объяснение этого явления в 1961 г. независимо друг от друга высказали Мери Лайон и Лиан Рассел. Они предположили, что одна из Х - хромосом в соматических клетках инактивируется случайным образом на ранних стадиях эмбрионального развития. Все клетки-потомки зародышевых клеток содержат такую же инактивированную Х - хромосому. М. Лайон проводила эксперименты с самками мыши, имеющих пятнистую окраску шерсти: в клетках одних пятен экспрессируется аллель одной Х - хромосомы, полученной от матери, а в клетках соседних пятен – аллель другой Х - хромосомы, полученной от отца. Такой фенотип обусловлен инактивацией разных Х - хромосом, несущих различные аллели этого гена. Сходная мозаика шерсти наблюдается у кошек. У котов все соматические клетки содержат одну Х - хромосому (от матери), то есть коты гемизиготны по этим генам и мозаичность по Х -сцепленным генам у них не обнаруживается. Кроме дальтонизма у человека встречается заболевание ангидротическая эктодермальная дисплазия, при котором у мужчин (гемизиготных по мутантному гену) отсутствуют зубы и потовые железы. У гетерозиготных женщин по этой мутации имеются участки кожи без потовых желёз. Это связано с инактивацией разных Х - хромосом, несущих либо нормальный, либо мутантный ген. Гипотеза Лайон не объясняла механизма инактивации хромосом у млекопитающих, но
вызвала научный интерес по дальнейшему изучению половых хромосом.
Объяснению явлений, связанных с Х – сцепленным наследованием, способствовало то, что долгое время Y - хромосома считалась генетически инертной. Затем были обнаружены гены в Y - хромосоме, передающиеся исключительно от отца к сыну и никогда к дочерям. К таким аномалиям относятся волосатые мочки ушей (hypertrichosis pinnae auris), волосатость всего тела, перепонка между пальцами ног. Признаки, передающиеся от отца к сыну исключительно по мужской линии, называются голандрическими. У человека в Y - хромосоме были обнаружены гены, детерминирующие развитие семенников, отвечающие за сперматогенез, контролирующие интенсивность роста тела, конечностей и зубов. К началу 2002 г. в Y - хромосоме картировали 35 генов, 7 из которых вызывают наследственные заболевания. К выше перечисленным заболеваниям добавились такие, как, пигментный ретинит, несколько форм азооспермии, дисхондростеоз, гондабластома и др. К настоящему времени установлено, что Y - хромосома имеет 78 генов, состоит из 50 млн. нуклеотидных пар, – это самая малая хромосома человека, и первая, полностью секвенированная Y - хромосома живого организма в рамках международного проекта «Геном человека». Интерес к Y - хромосоме вызван тем, что при некоторых морфологических дефектах её, можно проследить наследование такой хромосомы в потомках через очень большое число поколений. Так в вестнике ВОГиС появилась электронная версия статьи известного генетика И.Захарова, о наследовании Y - хромосомы потомками Чингисхана в Монголии и Центральной Азии. Их число превышает 16 млн. человек. Эти исследования были проведены группой учёных, возглавляемых оксфордским генетиком К. Тайлером Смитом, и опубликованы в авторитетном журнале по генетике человека (American J. Hum. Genet. 2003.V.72. P. 717-721). Такие исследования позволяют проследить пути миграции народов, установить родственные связи этнических групп, не связанных, казалось бы, ни территориальными, ни религиозными узами.
О значимости Y - хромосомы в определении мужского пола подтверждает тот факт, что при увеличении числа X - хромосом (XXY; XXXY; XXXXY), при наличии Y -хромосомыпол фенотипически проявляется как мужской. Известный медикам синдром Клайнфельтера у мужчин, сопровождающийся высоким ростом, евнухоидным и слегка феминизированным габитусом, плохим ростом бороды, маленькими тестикулами, гиалинозом тестикулярной ткани, гинекомастией, сниженным IQ, был объяснён цитологами в 1959г. анеуплоидией по X - хромосоме (XXY). Частота его проявления довольно высокая 1:700.Такой тип наслндования половых хромосом описан у собак и свиней. Лишняя Х – хромосома в генотипе у котов детерминирует у них развитие черепаховой окраской, что в норме встречается только у кошек. Реже встречаются варианты синдрома Клайнфельтера (XXXY – 1: 2500; XXXXY - очень редкий). Отсутствие Y -хромосомы при наличии хотя бы одной X -хромосомы (X 0) приводит к развитию женского фенотипа – синдром Шершевского – Тернера. Он встречается 1: 2500 женских особей и характеризуется тем, что индивидум имеет низкий рост, слабое развитие молочных желез, рудиментарные яичники, фиброзный тяж на месте гонад, «рыбий рот», «лицо сфинкса», крыловидные складки шеи. Этот синдром встречается у домашней мыши и козы. При триплоидии XXX - хромосом, встречающейся у женщин с частотой 1:1000, наблюдается лёгкая олигофрения, непостоянные нарушения функции гонад. Очень редко встречаются женщины с XXXX и XXXXX -типом. Обычно это физические нормальные женщины, имеющие тяжёлую умственную отсталость. В 1949г. М. Барр установил, что нормальные соматические клетки мужских и женских особей характеризуются наличием или отсутствием в них полового хроматина, названного в честь открывателя тельцем Барра. Приготовленные препараты из мазков слизистой оболочки рта, окрашиваются специальными красителями. У нормальных мужских особей тельце Бара отсутствует, нормальные женские особи имеют одно такое тельце. При синдроме Тернера - X 0, тельце Барра отсутствует. При синдроме Клайнфельтера – XXY, тельце Бара присутствует. В том случае, когда присутствует три или четыре X - хромосом, количества телец Барра всегда на единицу меньше числа X -хромосом. Определения количества телец Барра в соматических клетках позволяет выявить нарушение численности X - хромосом.
По голандрическому типу наследуются некоторые признаки у рыб. Например, у живородящей аквариумной рыбки гуппи один из признаков – тёмное пятно на спинном
плавнике обусловлено геном, локализованном в Y - хромосоме. Тёмное пятно передаётся только от отца к сыну, поэтому все самцы будут иметь его.
У аквариумных рыбок медаки (Oryzias latipes) красный цвет характерен для самцов, белый — для самок: доминантный ген R находится в Y -хромосоме, его рецессивный аллель r - в X -хромосоме
Р♀ XrXr x♂ XrYR
F1♀ XrXr x♂ XrYR
белые красные
Как же осуществляется наследование, сцепленное с полом, в том случае, когда гетерогаметный пол является женским, как, например, у кур, шелкопряда, некоторых видов рыб? У них самки несут XY -, а самцы XX -хромосомы. кур наследуется сцепленно с полом по типу крисс-кросс целый ряд признаков: полосатое оперение плимутроков, аспидная окраска ног. Так, поперечно-полосатое оперение обусловлено особым типом распределения окраса по перу и контролируется доминантным геном B, находящимся в Х -хромосоме. Например, полосатую курочку скрестили с черным петухом:
Р ♀ BY x ♂ bb
F1♂ B b: ♀ bY
пестрый черная
петух курица
5.2.3. Половой диморфизм и переопределение пола в онтогенезе
На ранних стадиях развития человеческий зародыш не имеет специфических признаков пола и считается потенциальным гермафродитом. К пятой неделе беременности зародышевые клетки формируют наружную корку (кортекс) и внутреннюю медуллярную пластинку. Впоследствии кортекс может дать начало яичникам, а медуллярная пластинка – семенникам. В это время эмбрион имеет недеференцированные Вольфовы (мужские) и Мюллеровы (женские) протоки.
Половая дифференциация эмбриона человека начинается с образования гонад. Так, у шестинедельного эмбриона, который имеет длину 12 мм, пол еще не различим, гонады внешне одинаковы. У зародыша к седьмой неделе внутриутробного развития, имеющего длину 13 мм, начинается развитие медуллярной пластинки, и появляются первые признаки мужских гонад — семенников. Гонады женского эмбриона в это время индифферентны. Процесс дифференциации яичников начинается в них на неделю позже. Одновременно происходит развитие соответствующей системы протоков, а другая – деградирует.
В 12-недельном возрасте пол эмбриона человека хорошо виден. Процесс дифференциации пола обусловлен гормонами, выделяемыми эндокринными железами, кортикулярным и медулярным слоями, а позднее — половыми железами. Развитие вторичных половых признаков идет под влиянием различных гормонов. Опыты М. М. Завадовского в 20-е годы показали, что кастрация самок птиц приводит к развитию у них признаков петуха и наоборот.
Главным действующим началом в дифференциации пола являются гены, контролирующие уровень гормональной секреции мужского или женского направления. При этом мужские гормоны (андрогены) и женские (прогестерон и эстроген) вызывают не только развитие вторичных половых признаков, но и соответственно маскулинизацию яичников (появление в них мужских половых клеток) и феминизацию семенников (появление в них женских половых клеток). При развитии зародыша по женскому типу на 12-ой неделе беременности в овогониях – клетках яичников - начинается мейоз и можно обнаружить первичные овоциты (овоциты I-го порядка). К 25-й неделе внутриутробного развития мейоз во всех овоцитах прекращается вплоть до пубертантного периода. То есть только через 10-15 лет все овоциты I-го порядка проходят сложный процесс мейоза с образованием овоцитов II-го порядка, редукцией хромосом, элиминацией первого и вторичных полярных телец и образованием полноценной яйцеклетки. Для неё характерно уменьшенное вдвое число хромосом и значительное количество запасных веществ. Самые крупные яйцеклетки в виде желтка яйца имеются у птиц. Они обеспечивают питание зародыша на протяжении всего эмбрионального развития. У млекопитающих развивающийся зародыш получает питательные вещества от материнского организма, поэтому их яйцеклетки значительно меньше. У особей мужского пола сперматоциты I-го порядка образуются только после достижения половозрелости. Каждая из них делится мейотически и даёт начало четырём клеткам – сперматидам с половинным числом хромосом. Без дальнейших делений ядра эти клетки превращаются в сперматозоиды. При слиянии яйцеклетки и сперматозоида образуется зигота, дающая начало новому организму.
Определение пола у растений затруднено тем, что они характеризуются большим разнообразием половых форм цветков:
Гермафродитны е (обоеполые), образующие тычинки и пестики -  ;
;
Тычиночны е (мужские, андроцейные), формируют только тычинки - ♂;
Пестичны е (женские, геницейные), формируют только пестики - ♀.
Половые типы древесных растений представлены следующими формами:
Обоеполые – растения с гермафродитными цветками: магнолиевые (Magnoliacea), ильмовые (Ulmaceae), липовые (Tiliaceae), розоцветные (Rosaceae), бобовые (Fabaceae), миртовые (Myrtaceae), рутовые (Rutaceae), кленовые (Aceracea (некоторые виды)
Раздельнополые однодомные – растения, как с пестичными, так и с тычиночными цветками (все хвойные (Pinopsida), платановые (Platanacaeae), самшитовые (Buxaceae), буковые (Fagaceae), березовые (Betulaceae), ореховые (Juglandaceae), кленовые (Aceracea: клён серебристый – Acer saccharinum);
Андроцейно-однодомные - растения, на которых образуются как тычиночные, так и обоеполые цветки (некоторые виды клёнов, каштан конский - Aesculus hippocastanum.
Геницейно-однодомные - растения с пестичными и обоеполыми цветками (некоторые виды клёнов, крушина – Frangula alnus).
Тримоноцейные – растения, на которых образуются цветки трёх типов: тычиночные, пестичные и обоеполые. – лимонник китайский - Schisandra chinensis, ясень обыкновенный – Fraxinus excelsior, клён остролистный – Acer platanoides, клён ложноплатановый – Acer pseudoplatanus, клён полевой - Acer campestre, клён татарский- Acer tataricum.
Раздельнополые, двудомные – а) андроцейные (мужские)- растения только с мужскими цветками; б) гинецейные (женские) – растения только с женскими цветками. (ивовые – Salicaceae), актинидия коломикта – Actinidia kolomicta, облепиха –Hippophae rhamnoides, клён ясенилистный - Acer negundo, ясень пенсильванский – Fraxinus pennsylvanica /
Кроме этого древесные растения различаются по типу опыления: ветроопыляемые и насекомоопыляемые растения. Выше перечисленные половые типы древесных растений не отображают всей сложности их сексуализации. Так клен остролистный и клён ложноплатановый могут быть представлены деверьями, на которых развиваются три половых типа цветков: тычиночные, пестичные и обоеполые. Или же только два: пестичные и обоеполые - у одних и тычиночные и обоеполые - у других. Всегда описываемый в дендрологической литературе лимонник китайский как тримонацейное растение в природе имеет раздельнополые, двудомные особи, в то время как актинидия коломикта вместо раздельнополых двудомных растений имеет особи, как с пестичными так и тычиночными цветками на одном растении (раздельнополые однодомные). Среди раздельнополых однодомных растений, к которым относится карельская берёза - Betula pendula Roth. var. karelica Merkl., встречаются особи мужской сексуализации (только андроцейные, мужские серёжки) так и женской сексуализации (только геницейные женские серёжки) (С.П. Погиба, 1988). Такие же особенности отмечает Р.Ф.Кудашева (1965) у лещины обыкновенной.
Это многообразие половых типов растений затрудняло восприятие понятия пола у растений. Сохранились фрески, на которых изображены ассирийские жрецы, опыляющие женские соцветия финиковой пальмы, следовательно, они знали, что у этого вида растений, как и у животных, имеется два пола. Несмотря на то, что арабы задолго до нашей эры практиковали искусственное опыление финиковых пальм, в Европе эти знания были утеряны. В 1759 г. Петербургская Академия наук объявила конкурс для выяснения этого вопроса. Премию в 1760 г. получил К.Линней за работу «Исследование пола у растений».
Описание микроспорогенеза и микрогаметогенеза лиственных и хвойных древесных растений, а также эмбриологическое развитие изложено в главе 4. Следует отметить, что, несмотря на общие положении, присущие голосеменным и покрытосеменным, каждый вид имеет определённые особенности эмбрионального развития.
На дифференциацию пола у высших растений значительное влияние оказывает действие растительных гормонов — ауксинов.
Еще в 1926 году С. С. Четвериков при проявлении признака, кроме генотипической и паратипической (экологической) среды, учитывал эмбриональное поле. (Погиба и др., 2004).
Наилучшим доказательством наследственной бисексуальности организма является изменение пола в онтогенезе в естественных или искусственных условиях.
У человека и млекопитающих, как правило, при развитии разнополых близнецов (две яйцеклетки оплодотворены двумя сперматозоидами) гормональное переопределение пола в эмбриогенезе затруднено тем, что дифференциация пола наступает раньше, чем начинается продуцирование соответствующих гормонов. Однако имеются случаи, когда изменение пола происходит в эмбриогенезе. Так, у крупного рогатого скота в случае рождения однополой двойни особи развиваются нормальными. Когда двойня разнополая, то бычки развиваются нормально, а телочки часто оказываются интерсексами: наружные гениталии женского типа, а внутренние органы — мужского. Такие животные бесплодны и названы фримартинами. Подобные изменения вызываются тем, что семенники мужского эмбрион иногда раньше начинают выделять мужские гормоны в кровь, которые и вызывают изменение женского эмбриона.
Классическим примером переопределения пола в природных условиях является изменение пола у морского червя — Boneleia viridis. Свободно плавающие личинки этого червя развиваются в самок. Личинки, прикрепляющиеся к хоботку самки, развиваются в самцов. При отделении от самки личинки превращаются в интерсексов. Как установлено, в хоботке самки морского червя имеются химические вещества, переопределяющие пол. Можно предположить, что такое определение пола является биологически выгодным для данного организма.
В последние годы внимание исследователей все более привлекает переопределение пола путем воздействия различными гормональными препаратами. У ряда животных удается получить полное преобразование пола, вплоть до способности формирования измененными особями половых клеток противоположного пола. Такие результаты получены у тритона, некоторых видов лягушек, рыб, птиц и ряда других животных, а также у растений и человека. Считается, что отношение полов 1:1 наиболее прогрессивно в эволюционном плане.
При рождении (вторичное отношение полов у человека) на 100 девочек приходится 103—106 мальчиков. В онтогенезе (третичное отношение полов) соотношение меняется:
Женщины Мужчины
18 лет 100 100
50 лет 100 95
57 лет 100 90
67 лет 100 70
87 лет 100 50
100 лет 100 21
Почему женщины более жизнеспособны в сравнении с мужчинами, остается неизвестным. (Н.Н. Дубинин, 1976)
У животных можно найти различные соотношения полов (100 самок и 0 самцов, 0 самок и 100 самцов). Есть виды (дрозофила, божьи коровки), имеющие однополые линии. Как показал Я. Я. Лусис (1947), это явление имеет приспособительное значение в жизни популяции.
Среди древесных растений встречаются однодомные и двудомные виды. При создании лесосеменных плантаций необходимо учитывать не только сексуализацию растений, но и изменение её с возрастом. Почти у всех хвойных молодые плантации испытывают недостаток пыльцы и требуют доопыления. Мужские стробилы в достаточном количестве развиваются позднее. При создании промышленных плантаций сортовой лещины обязательно с основными сортами планируют размещение сорта-опылители.
Кроссинговер
Понятие о кроссинговере
В предыдущей главе при изучении мейоза было показано, что в профазе I наблюдается явление перекреста гомологичных хромосом (хиазмы) и обмен участками у их хроматид. Этот процесс, открытый Г. Морганом на дрозофиле в 1911 году, получил название перекреста хромосом, или кроссинговера (от англ. crossing over – перекрест поверх). В результате кроссинговера нарушается сцепление генов, расположенных в одной хромосоме. Гаметы с хромосомами, претерпевшими кроссинговер, называются кроссоверными, а гаметы, образованные без кроссинговера, - некроссоверными. В соответствии с этим и особи, возникшие с участием кроссоверных гамет, называются кроссоверными или рекомбинантными, а образованные без них – некроссоверными или нерекомбинантными.
Явление кроссинговера было подтверждено не только на дрозофиле, но и в опытах с кукурузой Г. Крейтон и Б. Мак-Клинток (Г. В. Гуляев, 1971; И. Ф. Жимулев, 2007). Пример явления кроссинговера описан Г. В. Гуляевым при скрещивании двух линий кукурузы, хорошо различающихся по окраске эндосперма и консистенции алейронового слоя. Одна линия имела в гомозиготном состоянии доминантные гены C и S, контролирующие образование окрашенного эндосперма и гладкого алейрона, а другая – их рецессивные аллели с и s, определяющие развитие неокрашенного эндосперма и морщинистого алейрона. Гибридные растения этого скрещивания имели окрашенный эндосперм и гладкий алейрон.
Такие дигетерозиготные растения (CcSs) при независимом комбинировании генов должны были бы образовать в равном количестве четыре типа гамет: CS, Cs, cS, cs. При анализирующем скрещивании с отцовской рецессивной гомозиготной линией (ccss) можно было бы ожидать расщепления в отношении 1 C - S (окрашенные гладкие): 1 С- ss (окрашенные морщинистые): 1 сс- S (неокрашенные гладкие): 1 сс ss (неокрашенные морщинистые). В действительности же 96,4% всех зерен имели признаки, свойственные исходным родительским линиям (48,2% окрашенных гладких и 48,2% неокрашенных морщинистых), и только 3,6% зерен были с новым сочетанием признаков.
Объяснение полученных результатов возможно при допущении сцепления генов. Гены C и S находятся в одной хромосоме, они сцеплены и поэтому во время мейоза попадают в одну гамету, их же рецессивные аллели a и s, сцепленные в другой хромосоме, попадают в другую гамету. Так образуется более 96% некроссоверных гамет и соответственно 96%некроссоверных зерен. Однако сцепление между генами C и S и c и s не полное. При конъюгации хромосом, несущих эти пары генов, происходит кроссинговер, в результате которого доминантные и рецессивные аллели обмениваются местами, попадая из одной гомологичной хромосомы в другую и образуя кроссоверные гаметы Cs и cS. В данном случае образовалось 3,6 % кроссоверных гамет и соответственно такое же число кроссоверных, или рекомбинантных зерен.
Таким образом, кроссинговер способствует разделению сцепленных генов, и тем самым увеличивает разнообразие наследственного материала.






