У покрытосеменных растений женский гаметофит называется зародышевым мешком, который закладывается и развивается внутри семяпочки. Развитию женского гаметофита предшествует макроспорогенез.
В субэпидермальном слое клеток нуцеллуса молодой семяпочки обособляется археспориальная клетка, чаще только одна. Клетка археспория растет, превращаясь в материнскую клетку макроспор.
В результате двух делений мейоза в материнской клетке образуется тетрада макроспор, расположенных обычно линейно. Каждая из клеток тетрады по числу хромосом является гаплоидной. Чаще всего только одна из клеток, а именно базальная, продолжает развитие, остальные три дегенерируют.
В результате трех эквационных делений ядра макроспоры в зародышевом мешке возникает восемь наследственно одинаковых ядер. Ядра занимают полярное положение:
- четыре из них оказываются лежащими ближе к микропиле (место проникновения пыльцевой трубки),
- а четыре других - в противоположном конце зародышевого мешка, называемым халазальным.
Из четырех ядер, располагающихся в микропилярном полюсе зародышевого мешка, три окружаются слоем цитоплазмы и обособляются в отдельные самостоятельные клетки, образуя яйцевой аппарат, состоящий из яйцеклетки и двух синергид. После оплодотворения развивается в норме только одна, а две другие разрушаются.
Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального полюса. Слившиеся в центральной части зародышевого мешка два гаплоидных ядра образуют одно диплоидное - вторичное, или центральное, ядро. Это ядро с цитоплазмой зародышевого мешка называют центральной клеткой.
Оставшиеся в халазальном конце зародышевого мешкатри ядра также обособляются в клетки, называемые антиподами (рис. 4.8).
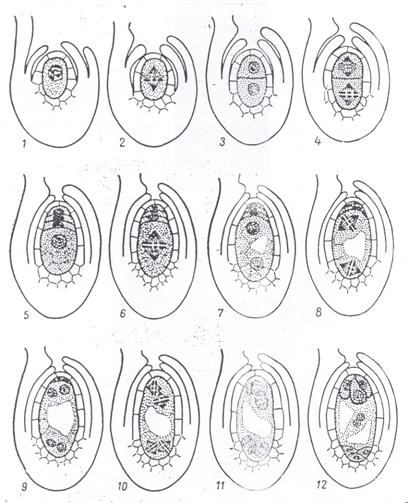
Рис. 4.8. Схема макроспорогенеза (1-5) и макрогаметогенеза (6-12) у покрытосеменных растений (М. Е. Лобашев. 1969).
4.4.4. Эндоспермогнез*)
Эндосперм, или ткань, питающая зародыш в семенах хвойных растений развивается непосредственно из клеток женского гаметофита. В этом состоит главное различие между эндоспермом голосеменных и покрытосеменных растений, у большинства из которых эндосперм образуется из центральной клетки зародышевого мешка после слияния ядра этой клетки с ядром спермия. Интенсивный рост женского гаметофита и отложение в его клетках запасных питательных веществ начинается сразу после оплодотворения яйцеклетки.
На ранних стадиях развития зародыша центральная часть женского гаметофита разрушается, образуя полость, в которую проникает зародыш после выхода из архегония. Роль клеток зародыша, при этом, как полагают, не пассивная. Они могут выделять энзимы, необходимые для переваривания содержимого клеток женского гаметофита, и активно абсорбируют продукты лизиса. Полость в гаметофите, образующаяся путем энзиматического действия, получила название коррозионной или зародышевого канала.
Резервные вещества эндосперма семян хвойных представлены запасными белками, жиром и крахмалом. В период роста зародыша в эндосперме хвойных четко различаются две зоны: внутренняя зона клеток с крахмалом и широкая внешняя зона, клетки которой наиболее богаты запасными веществами. Внутренняя зона крахмалоносных клеток полностью абсорбируется в период роста зародыша и в зрелых семенах хвойных эндосперм состоит из однородных по строению и содержанию запасных веществ клеток.
При образовании эндосперма в семенах покрытосеменных растений чаще всего наблюдается следующая картина: верхнее и нижнее полярные ядра в зародышевом мешке сливаются, образуя вторичное, или центральное ядро, которое затем сливается с одним из спермиев, входящих в зародышевый мешок. Возникает первичное ядро эндосперма, которое начинает делиться после более или менее продолжительного периода покоя. Стимул к росту эндосперм получает после оплодотворения яйцеклетки. Эндосперм покрытосеменных с момента слияния спермия с центральным ядром зародышевого мешка является триплоидным.
*) Разделы 4.4.4 и 4.4.5 подготовлены В. В. Трениным (2000).
Считается, что именно вследствие того, что эндосперм покрытосеменных возникает в результате оплодотворения и имеет двойственную наследственность, он отличается большей активностью роста и получает преимущество перед другими тканями семяпочки в поддержании жизни зародыша на самых ранних стадиях развития. Возникновение в ходе эволюции растений гибридного эндосперма повысило приспособляемость к внешним условиям и явилось одним из условий широкого распространения покрытосеменных в современную геологическую эпоху. Первичное ядро эндосперма обычно начинает делиться раньше, чем оплодотворенное ядро яйцеклетки.
По особенностям возникновения и раннего развития различаются три основных типа эндосперма покрытосеменных растений: ядерный или нуклеарный, клеточный или целлюлярный и базальный. Независимо от способа возникновения эндосперм всех типов впоследствии становится клеточным. В зрелых семенах у одних видов покрытосеменных эндосперм более или менее хорошо развит, у других он вовсе не образуется, или, образовавшись, рано поглощается развивающимся зародышем. В семенах без эндосперма запасные питательные вещества откладываются в самом зародыше, в его семядолях.
Ядерный тип эндосперма характеризуется тем, что в нем подобно эндосперму хвойных, образованию клеточных стенок предшествует период свободно ядерных делений. Число свободных ядер у разных видов варьирует от 4-8 до 4000. Формирование клеточных стенок при ядерном эндосперме может начинаться с периферии зародышевого мешка и идти к центру, либо с верхнего (микропилярного) конца его и идти к нижнему. В одних случаях клетки эндосперма заполняют всю полость зародышевого мешка, в других - не всю, оставляя свободной обычно центральную полость.
Клеточный тип эндосперма покрытосеменных характеризуется тем, что здесь после первого и каждого из последующих делений ядер эндосперма сразу же образуются клеточные оболочки. Базальный тип эндосперма является промежуточным между ядерным и клеточным типами. Он отличается тем, что после первого деления ядра эндосперма образуется клеточная перегородка, которая делит зародышевый мешок на две неравные части: большую верхнюю и меньшую нижнюю. В верхней части эндосперм сначала образует ряд свободных ядер, и только позднее между ними образуются клеточные стенки. В нижней части либо также сначала образуется некоторое число свободных ядер, либо сразу же образуются клетки.
В период образования клеток в эндосперме деление ядер опережает формирование клеточных стенок. Благодаря этому в эндосперме часто образуются многоядерные клетки; ядра в тканях клеток нередко сливаются, становясь округлыми, часто причудливой формы. Подобное явление описано и для эндосперма семян хвойных на ранних фазах клеточной стадии.
4.4.5. Эмбриогенез*)
По анатомическим особенностям эмбриогенеза как голосеменные, так и покрытосеменные являются очень неоднородными группами растений. В развитии зародыша хвойных, как и всех голосеменных растений принято выделять три основные фазы: 1 - развитие проэмбрио (проэмбриогенез), 2 - ранний эмбриогенез, 3 - поздний эмбриогенез. Фаза проэмбриогенеза включает стадии с момента оплодотворения (слияния ядер гамет) и до начала удлинения суспензора при выходе зародыша из архегония. Первое деление ядра зиготы у хвойных проходит внутри архегония и в подавляющем большинстве случаев не сопровождается образованием клеточных перегородок. После второго деления у сосновых свободные ядра перемещаются в основание архегония, располагаются в одной плоскости и синхронно делятся. В результате образуются два яруса, или этажа клеток, между которыми формируется клеточная перегородка. Клетки верхнего яруса со стороны цитоплазмы архегония клеточной стенки не имеют. В дальнейшем клетки верхнего и нижнего ярусов могут делиться синхронно, но чаще синхронное деление наблюдается только в верхнем ярусе. У сосновых в развитии проэмбрио наблюдается два свободноядерных деления. У других хвойных их может быть от двух до пяти. У большинства хвойных, исключая сосновые, в нижнем ярусе насчитывается более четырех клеток и расположены они неупорядоченно.
Фаза раннего эмбриогенеза у хвойных начинается удлинением суспензора и заканчивается заложением в теле зародыша меристемы корня. Разные виды хвойных значительно отличаются по особенностям раннего эмбриогенеза. Зародыш в фазе раннего эмбриогенеза имеет шаровидную или удлиненно-цилиндрическую форму. Между полюсами зародыша в этой фазе эмбриогенеза нет резкой границы. Клетки зародыша слабо дифференцированы и характеризуются меристематической организацией, то есть они округлые по форме, имеют крупные ядра, а в густой цитоплазме нет больших вакуолей.
Очень часто в фазе раннего эмбриогенеза в коррозионной полости эндосперма наблюдается несколько зародышей. Полиэмбриония, или многозародышевость, у хвойных растений может быть следствием оплодотворения нескольких яйцеклеток из разных архегониев одного женского гаметофита (простая, или архегониальная полиэмбриония) или следствием расщепления зародыша на несколько самостоятельно развивающихся зародышей (расщепленная или кливажная полиэмбриония), или и того и другого вместе.
В конце фазы раннего эмбриогенеза зародыш принимает булавовидную форму с правильной округлой вершиной. Деления часто наблюдаются в клетках, прилегающих к суспензору, но образующиеся новые клетки больше к суспензору не добавляются и образуют массу упорядоченно расположенных клеток, ориентированных в поперечном к оси зародыша плане. В дальнейшем эти клетки становятся клетками корневого чехлика. Корневой чехлик зародыша хвойных очень массивный. В конце фазы раннего эмбриогенеза зародыш большинства хвойных более чем на половину своей длины состоит из клеток будущего корневого чехлика.
В фазе позднего эмбриогенеза у хвойных происходит активный гистогенез (формирование тканей) и рост зародыша. Зона клеток, ориентированных в поперечном плане, делит зародыш на микропилярную часть, где формируется корневой чехлик, и халазальную, где идет развитие оси гипокотиль-побег. Зрелый зародыш хвойных несет несколько семядолей и дифференцирован на две полярные меристемы побега и корня. Гистогенез в зародыше в фазе позднего эмбриогенеза сопровождается синтезом и накоплением в клетках всего зародыша запасных веществ. Поздний эмбриогенез заканчивается переходом зародыша в покоящееся состояние.
Размеры клеток в ходе эмбриогенеза у хвойных прогрессивно уменьшаются. Цитохимическое изучение ДНК в раннем эмбриогенезе показало, что в первых делениях проэмбрио количество ДНК, приходящееся на ядро, кратно уменьшается, достигая уровня 16 С на четырехядерной стадии и 4 С на стадии 16-клеточного проэмбрио. (Буквой С обозначается условное количество ДНК, имеющееся в гаплоидном ядре клетки данного вида). Предполагается, что накопленная в ядре яйцеклетки ДНК последовательно распределяется между дочерними ядрами в ходе первых делений, которые происходят без фазы синтеза ДНК. Все или подавляющее большинство ядер клеток зародыша в период перехода к состоянию покоя блокируются на стадии G1 клеточного цикла.
Зрелый зародыш в семени хвойных окружен эндоспермом и защищен семенной кожурой. Семенная кожура развивается из интегумента и тканей, расположенных в халазальной части семяпочки. Таким образом, семена хвойных имеют тройственную природу: семенная кожура является частью старого спорофитного поколения, эндосперм - женского гаметофита, а сам зародыш - это новое спорофитное поколение.
Электронно-микроскопическое исследование эмбриогенеза позволило установить природу цитоплазменных органелл в клетках зародыша растений. Для хвойных большая заслуга в этом принадлежит французскому ученому А.Камефору. За период с 1957 по 1971 год А.Камефор опубликовал около 30 статей, в которых на примере нескольких видов хвойных и гинкго проследил изменения в ультраструктуре формирующейся яйцеклетки и проэмбрио, что позволило ему разработать концепцию так называемой неоцитоплазмы (Camefort, 1968). Неоцитоплазма, или новая цитоплазма образуется в период оплодотворения и первых свободноядерных делений проэмбрио. Неоцитоплазма появляется и существует внутри архегония, вокруг слившихся в ходе оплодотворения ядер мужской и женской гамет. Цитоплазма клеток зародыша целиком формируется из неоцитоплазмы. Все пластиды, часть митохондрий и рибосом наследуются зародышем от спермия. Яйцеклетка привносит свою часть рибосом и митохондрий, причем у некоторых видов хвойных это - наибольшая часть митохондриона неоцитоплазмы.
Пластиды яйцеклетки в построении цитоплазмы зародыша не участвуют. В этом отношении хвойные кардинально отличаются от покрытосеменных, у большинства которых наблюдается однородительское материнское наследование пластид.
Более 100 лет оставалась загадочной природа крупных включений: в яйцеклетке хвойных, так называемых телец Гофмейстера (рис. 4.9). Только с помощью электронного микроскопа удалось установить, что они образованы мембранами гипертрофированных пластид. В.В.Тренин (1984) на основании собственных данных и анализе литературы сделал вывод о том, что наличие подобных образований в яйцеклетке голосеменных есть специфическая для этих растений форма изоляции той части цитоплазмы яйцеклетки, которая не участвует в построении новой цитоплазмы зародыша.

Рис. 4.9. Схема строения зрелого архегония сосновых (Тренин В. В., 2000).
В результате последовательного ряда делений зиготы возникают недифференцированные многоклеточные образования. По мере дальнейшего развития По особенностям эмбриогенеза покрытосеменные растения оказались еще более неоднородной группой, чем хвойные. Общепринятой классификации типов развития зародыша у покрытосеменных нет. Обычно во вполне сформированном виде, еще до прорастания семени, зародыш покрытосеменных имеет семядоли, подсемядольное колено (гипокотиль), почечку с зачатками стебля и листьев, зачаток корня и корневой чехлик.
По количеству семядолей у зародыша и проростков покрытосеменные делятся на два класса - однодольные и двудольные. Наиболее часто встречается зародыш с двумя семядолями. Первые стадии развития зародыша проходят одинаково в обоих этих классах покрытосеменных растений.
Олодотворенная яйцеклетка, которую называют зиготой, начинает делиться после более или менее продолжительного периода покоя. В результате первого деления образуются две неравные клетки: менее крупная, называемая апикальной, или терминальной, и более крупная, или базальная. Из базальной обычно образуется подвесок или суспензор, а из апикальной - основные части зародыша у его основания около подвеска дифференцируется гипофизис, дающий начало инициальным клеткам коры и чехлика корня, а на верхушечке его против гипофизиса дифференцируется эпифизис, дающий начало инициальным клеткам эпидермиса и коры зачатка стебля. Затем в виде бугорков начинается заложение семядолей.
Одновременно с наружной у зародыша происходит и внутренняя дифференциация, которая заключается в образовании двух зародышевых тканей: наружной или туники и внутренней, корпуса. Из туники впоследствии развивается эпидермис, а из корпуса - кора и центральный цилиндр взрослого растения.
Приведенное описание является общей схемой развития зародыша покрытосеменных. У разных представителей покрытосеменых в развитии и строении зародыша наблюдаются многочисленные отличия от общей схемы. Как и у голосеменных растений, в семенах покрытосеменных может развиться несколько зародышей. Полиэмбриония у покрытосеменных возникает различными путями и может быть основана на базе, как полового, так и бесполого размножения (апомиксиса).
Вопросы для самопроверки
1. Назовите основные компоненты растительной клетки.
2. Назовите органеллы, видимые в световом микроскопе и те, которые различимы только в электронном микроскопе.
3. Назовите характерные особенности строения растительной клетки, отличающие ее от клеток животных.
4. Каковы основные функции пластид и вакуолей?
5. Назовите периоды клеточного цикла.
6. Опишите фазы митоза.
7. В каких органах растений проходит мейоз и из каких последовательных фаз он состоит?
8. В чем состоит основное биологическое значение митоза и мейоза?
9. Назовите основные эмбриологические процессы у высших растений.
10. В чем состоят главные отличия в эмбриологических процессах у хвойных и покрытосеменных растений?
11.Что такое двойное оплодотворение, его общебиологическое значение?
12. Какова природа эндосперма у голосеменных и покрытосеменных растений?
13.Что такое простая полиэмбриония в развивающихся семенах хвойных растений?
Глава 5.





