Краткая классификация генов
Из всего вышеизложенного можно видеть, что ген - это участок молекулы ДНК, контролирующий определенный признак организма. Или (по С.М. Гершензон 1979) - участок молекулы геномной (из гаплоидного набора) нуклеиновой кислоты, характеризуемый специфической для него последовательностью нуклеотидов, представляющий единицу функции, отличной от функции других генов и способный изменяться путем мутирования.
Согласно схеме С. Бензера, генетический материал разделяется на цистроны - единицы функции, мутоны - единицы мутации и реконы - единицы рекомбинации. Все эти единицы характеризуются различным количеством пар нуклеотидов. Цистрон отвечает за синтез полипептидной нити и может содержать до 1000 нуклеотидов.
Гены делятся на две категории:
- Структурные, или смысловые. Несут информацию о последовательности аминокислот в белковой молекуле, то есть определяют структуру белков. Комплекс структурных генов с геном-оператором составляют оперон.
- Акцепторные, или регуляторные гены – последовательности нуклеотидов, не имеющие кодирующих функций, но с помощью присоединения к ним разных белковых факторов, управляющие работой структурных генов. То есть, они являются местами специфического присоединения белков-репрессоров и белков-активаторов (С. М. Гершензон, 1983; М. Сингер, П. Берг, 1998).
К акцепторным генам относятся ген - оператор, ген - промотор, ген - терминатор.
Ген-оператор координирует проявление соседних генов, составляющих оперон. Оперон – это функциональная генетическая единица с размером, средним между размером гена и хромосомы. Оперон представляет совокупность совместно транскрибируемых генов, обычно контролирующих родственные биохимические функции. Он составлен из ряда линейно расположенных генов, структурная активность которых координируется прилегающим к ним регуляторным геном (рис. 6.16).
Ген-промотор – это стартовые точки на ДНК, к которым присоединяются РНК-полимеразы с тем, чтобы начать транскрипцию (согласно данным молекулярной генетики, начало транскрипции связано с присоединением к определенным последовательностям ДНК молекул РНК-полимеразы).
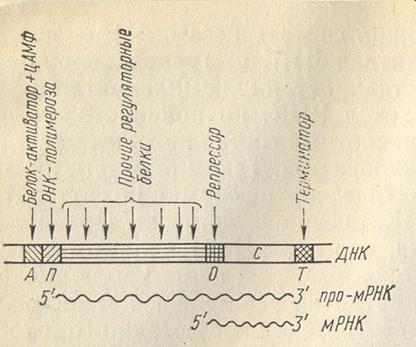
Рис. 6.16. Схема строения оперона у эукариот (Г. П. Георгиев, по С. М. Гершензон, 1983). А – участок, к которому прикрепляется белок-активатор; П – промотор; О – оператор; С – структурный ген; цАМФ – побочный продукт цепи синтеза пуринов, который конститутивно образуется особым ферментом из аденозинтрифосфата (АТФ).
Ген-терминатор – ген, прекращающий определенные действия других генов.
Выделяют также гены-супрессоры (подавляющие действие других генов), гены развития, стерильности, олигогены, полигены, гены, сцепленные с полом, гены, обусловливающие проявление тех или иных признаков организма и др.
Генетическая номенклатура
Впервые буквенную символику для обозначения признаков ввел Г. Мендель. По мере развития генетических знаний появилась необходимость в установлении названий исследуемых генов, обусловливающих проявление тех или иных признаков. Общие правила генетической символики впервые были предложены Комитетом по генетическим формам и номенклатуре Американского общества натуралистов в 1920 году. Совершенствование генетической номенклатуры продолжается до настоящего времени (И. А. Захаров, 1979; С. М. Гершензон, 1983; И. А. Захаров, Б. П. Мацелюх, 1986 и др.). При этом рекомендуется соблюдение ряда требований генетической номенклатуры. Некоторые из этих рекомендаций приведены ниже.
- При записи скрещиваний генотип женского родителя пишется первым.
2. В качестве стандарта используется так называемый дикий тип. При этом в качестве дикого типа принимается действительно дикая форма, предок культурных форм или лабораторных линий, или наиболее примитивная из культурных форм.
3. В качестве названий генов используются слова, характеризующие основное проявление гена. Это или одно слово, или комбинация нескольких слов, характеризующих эффекты данного гена. Например, известны гены томатов anthocyaninless (безантоциановый), without anthocyanin (без антоциана), yellow seedlings (желтые проростки).
4. Для обозначения генов рекомендуется использовать прилагательные или существительные латинского или английского языка, однако могут использоваться слова и из других языков.
5. Для генов, имеющих одинаковое или сходное проявление, но расположенных в разных локусах, принято использовать одинаковое название, но с цифровыми индексами. Так, гены томатов, вызывающие в мутантном состоянии асинапсис хромосом в мейозе, обозначены asynaptic 1, asynaptic 2 и т.д.
6. Название генов принято сокращать. Название сокращается до 1–4 букв (символ гена), причем берется такое минимальное количество букв, чтобы сокращенное обозначение не совпадало с любым другим для генов данного вида организмов. При сокращении названия гена, как правило, в качестве первой берется первая буква названия и к ней, если необходимо, добавляются непосредственно следующие за ней буквы или буквы из середины или конца слова, или буквы из других слов, входящих в название. Например: anthocyaninless – a; apricot – at; cabbage – cb. Для генов со сходным проявлением цифровой индекс приписывается и к их символу: as1 (asynaptic 1), as2 (иногда as1, as2). В публикациях символы генов набираются обычно курсивом.
7. Каждый ген представлен, по крайней мере, 2 аллельными состояниями, одно из них считается аллелью дикого типа. Аллель дикого типа обозначается символом с индексом «+»: sp+ или I+. В первом случае мутантный аллель sp, обозначаемый строчными буквами рецессивен по отношению к дикому типу; во втором, I, обозначаемый заглавной буквой – доминантный.
8. В качестве индексов используются сокращенные названия признаков, характерных для проявления данного мутантного аллеля: w (white) – аллель, вызывающий белую окраску глаз, wa (white-apricot) – розовая окраска глаз, wch (white-cherry) – вишневая окраска. Применяются также и другие индексы. Символы аллелей генов, чьи эффекты выявляются электрофоретически, все пишутся как доминантные, с заглавной буквы; так, аллели гена алкогольдегидрогеназы обозначены AdhF, AdhS, Adhn1 и т.п. Используется и более ранняя система символов для серии множественных аллелей вида: AY, AI, A, a, где дикий тип обозначается заглавной буквой без индексов и без знака плюс, рецессивные аллели – строчными буквами, доминантные –заглавными с соответствующими индексами.
9. При обозначении сложных генотипов рекомендуется следовать порядку, принятому для дрозофилы - вначале писать символы генов первой хромосомы, затем второй и т.д., а внутри группы сцепленных генов размещать их в порядке их локализации, отделяя промежутками без запятой. Гены разных групп сцепления разделяют точкой с запятой. Так, следующим образом будет записан гаплоидный генотип гамет дрозофилы: y, ct, v; ss, где символы y, ct, v относятся к генам I группы сцепления, ss – к III. Группы сцепления нумеруются в порядке их выявления, при этом используются римские цифры; когда завершается работа по идентификации групп сцепления с определенными хромосомами, для тех и других вводится единая нумерация (исходя из номеров хромосом). Пока работа по идентификации не завершена, возможны обозначения вида 2(I), относящиеся к группе сцепления I, идентифицированной с хромосомой 2.
10. Существуют три способа изображения генотипов диплоидов:
1) Aa Bb cc;
2) A/a B/b c/c, или +/a +/b c/c;
3) ABc/abc, или ++c/abc. Третий способ дает возможность наглядно изображать сцепление между генами: ++с/abc – все три гена сцеплены; ++/ab c/c – сцеплены только два первых гена.
11. Хромосомы принято нумеровать согласно убыванию их длин, измеряемых цитологически, причем номер 1 получает самая длинная хромосома (нумерация хромосом дрозофилы является исключением из этого правила). Для обозначения хромосом употребляются арабские цифры. Символы L и R обозначают левое и правое (условно) плечи хромосомы (например, 2L, 2R), а p и q – короткое и длинное плечи (например, 2p, 2q).
12. Для обозначения основных типов хромосомных перестроек используют следующие символы. Транслокации обозначаются символом Т. В скобках за ним указываются номера хромосом, вовлеченных в транслокацию: Т (1–2). Для инверсий, дупликаций и нехваток применяются соответственно символы In, Dp, Df, а в скобках указывается номер хромосомы с соответствующей перестройкой: In (1), Df (2). Для кольцевых хромосом используется символ R.
13. Особых правил для обозначения цитоплазматических генов нет. Для их сокращенных обозначений имелась тенденция использовать буквы греческого алфавита, хотя такие буквы иногда, в генетике грибов, используются и для обозначения ядерных генов. Сейчас предпочитают пользоваться символами, аналогичными символам ядерных генов. В записях генотипов рекомендовано символы цитоплазматических генов заключать в квадратные скобки [ p ] и размещать перед символами, относящимися к ядерным генам.
Транскрипция и трансляция
Реализация генетической информации, заключенной в последовательности нуклеотидов ДНК, включает две стадии.
На первой каждый ген служит матрицей для синтеза молекул РНК. На информационную РНК (иРНК) переписывается последовательность нуклеотидов определенного гена. Следовательно, в РНК закодирована последовательность аминокислот. Процесс переписывания или транскрипции происходит в ядре на одной из нитей ДНК. Транскрипция - это переписывание последовательности нуклеотидов гена с ДНК на иРНК. Далее, на второй стадии, иРНК перемещается в цитоплазму, где последовательность нуклеотидов переводится в последовательность аминокислот полипептида (белка). Этот процесс назван трансляцией. То есть трансляция - это перевод последовательности нуклеотидов гена в последовательность аминокислот белка.
Информация о синтезе белка зашифрована в последовательность нуклеотидов. Три нуклеотида, кодирующих одну аминокислоту в ДНК называются кодоногеном, а в иРНК - кодоном. Каждый кодоноген присоединяет комплементарный кодон. В процессе трансляции в рибосомах к кодонам иРНК подходят транспортные РНК (тРНК) с соответствующими антикодонами. Аминокислота, принесенная тРНК, только в том случае идет на синтез белка, когда триплет иРНК комплементарен триплету тРНК. (рис. 6.16).
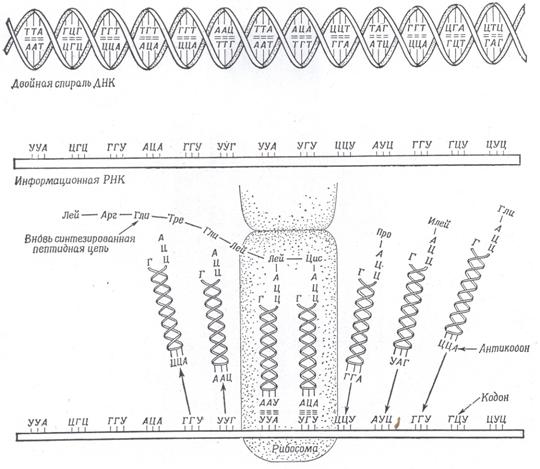
Рис. 6.16. Схема гипотетического механизма синтеза белка на рибосоме, отражающая отношения между триплетами спирали ДНК, комплементарными им триплетами информационной РНК и комплементарными последним триплетами (антикодонами) транспортных РНК. Молекулы транспортной РНК с присоединенными к ним специфическими аминокислотами приходят справа, занимают надлежащие места на информационной РНК у рибосомы. Переносят аминокислоты на растущую пептидную цепь, а затем (слева) отделяются от рибосомы и могут снова присоединять аминокислоты для дальнейшего синтеза белка. Растущая полипептидная цепь остается прикрепленной к одной и той же рибосоме (К. Вили,1968).
Процессы транскрипции и трансляции в клетках эукариот проходят значительно сложнее в сравнении с бактериальными.
Следует также отметить, что последующее изучение генов обнаружило их неоднородную структуру. Не все участки генов участвуют в кодировании аминокислот. Одни из них кодируют, как описано выше, а другие нет. Участки, способные кодировать белки называют экзонами, а участки ДНК, которые не участвуют в этом процессе – интронами. Чтобы не было разрывов в структуре белков, РНК после транскрипции, но перед процессом трансляции в результате работы специфических ферментов подвергаются так называемому сплайсингу. В результате сплайсинга отбрасываются некодирующие белки интроны, а экзоны соединяются в том порядке, в каком они были в ДНК (рис. 6.17).
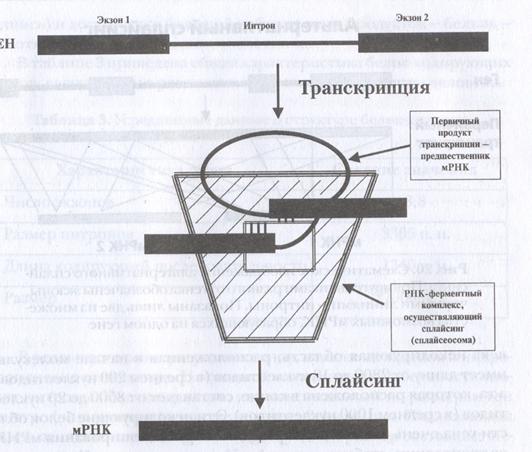
Рис. 6.17. Образование молекул мРНК на РНК-предшественнике в результате сплайсинга. Из РНК-предшественника вырезаются фрагменты, синтезированные с интронных последовательностей гена, а участки, синтезированные на экзонах, соединяются друг с другом, что в конечном итоге приводит к формированию зрелой функционально активной мРНК (В. З. Тарантул, 2003).
На первых этапах развития учения о молекулах информационных РНК, Ф. Крик выдвинул идею (в 1958 году) о главных процессах, идущих при синтезе белков, назвав её центральной догмой молекулярной биологии. Согласно центральной догме ДНК обладает способностью строить на генах молекулы информационной РНК (иРНК). Молекулы иРНК переходят в цитоплазму и передают код гена на синтез полипептидов (трансляция). Согласно догме эти процессы не имеют обратной связи. То есть полипептид не способен транскрибировать молекулы иРНК, а молекулы иРНК не способны транскрибировать молекулы ДНК. К настоящему времени выявлены процессы, которые не согласуются с центральной догмой молекулярной биологии (М. Д. Голубовский, 2000).
Вопросы для самопроверки
1. Как было доказано, что именно ДНК, а не белок, является носителем наследственности?
2. Из каких компонентов состоят нуклеотиды нуклеиновых кислот? Как они расположены?
3. Чем отличаются пурины от пиримидинов и какие азотистые основания входят в ту или иную группу?.чем отличается ДНК от РНК?
4. Охарактеризуйте модель структуры ДНК, предложенной Д. Уотсоном и Ф. Криком.
5. Чем обусловлено правило Э. Чарграффа и в чем оно заключается? Что такое коэффициент специфичности ДНК?
6. Как происходит репликация нуклеиновых кислот?
7. Какие из важнейших аминокислот входят в состав белка? Сколько их? Сколько аминокислот теоретически могут кодировать четыре типа нуклеотидов?
8. Что представляет собой генетический код? Покажите его в виде таблицы и в виде круга.
9. Каковы особенности генетического кода?
10. Приведите классификацию генов. На какие категории они делятся?
11. Какими рекомендациями пользуются при наименовании генов?
12. Что такое транскрипция и трансляция? Как происходит синтез белка у бактерий?
13. Что такое экзоны, интроны и сплайсинг?
14. В чем заключается центральная догма молекулярной биологии?
Глава 7.






