Транспорт веществ в бактериальную клетку. Существует два типа переноса веществ в бактериальную клетку: пассивный и активный. При пассивном переносе вещество проникает в клетку только по градиенту концентрации. Затраты энергии при этом не происходит. Различают две разновидности пассивного переноса: простую диффузию и облегченную диффузию.
Простая диффузия – неспецифическое проникновение веществ в клетку, при этом решающее значение имеет величина молекул и липофильность. Скорость переноса не значительна. Облегченная диффузия протекает с участиембелка-переносчика.Скорость этого способа переноса зависит от концентрации вещества в наружном слое.
Активный транспорт веществ в клетку. При активном переносе вещество проникает в клетку против градиента концентрации при помощи белка-переносчика – пермеазы. При этом происходит затрата энергии. Имеется два механизма активного транспорта.
Механизмы активного транспорта веществ в клетку. Активный транспорт: небольшие молекулы (аминокислоты, сахара) «накачиваются» в клетку, создается концентрация в 100-1000 раз выше, чем снаружи клетки
Второй механизм – транслокация радикалов,
обеспечивает транспорт сахаров (глюкоза, фруктоза). В процессе переноса происходит химическая модифиция – фосфорилирование. Для осуществления активного транспорта в бактерии имеется специальная фосфотрансферная система.
Фосфотрансферная система. Составной частью этой системы является активный фосфорилированныйбелок-переносчик(АФБ). АФБ связывает свободный сахар на наружной поверхности ЦПМ, транспортирует его в цитоплазму, где сахар освобождается в виде фосфата. Далее фосфаты вступают в цепь биохимических реакций, в результате образуются АТФ и ингредиенты для биосинтеза компонентов клетки. Это называется конструктивным метаболизмом.
Энергетический метаболизм. Типы метаболизма: окислитель-
ный, или дыхание; бродильный, или ферментативный; смешанный. Пути расщепления глюкозы при использовании ее в качестве ис-
точника углерода и энергии:
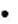 через образованиефруктозо-1,6-дифосфата(ФДФ-путь).Выход: 2 молекулы пирувата, 2 молекулы АТФ, 2 молекулы восстановленного НАД;
через образованиефруктозо-1,6-дифосфата(ФДФ-путь).Выход: 2 молекулы пирувата, 2 молекулы АТФ, 2 молекулы восстановленного НАД;
через пентозофосфатный путь(ПФ-путь).Выход: 1 молекула3-фосфоглицеринальдегида,3 молекулы СО2, 2 молекулы восстановленного НАД.
через путьЭтнера-Дудорова.Выход: 1 молекула АТФ, 1 молекула восстановленного НАД, 1 молекула восстановленного НАДФ.
Основные принципы культивирования микроорганизмов на питательных средах.
1.Использование всех необходимых для соответствующих микробов питательных компонентов.
2.Оптимальные температура, рН, концентрация ионов, степень насыщения кислородом, газовый состав и давление.
Микроорганизмы культивируют на питательных средах при оптимальной температуре в термостатах, обеспечивающих условия инкубации.
По температурному оптимуму роста выделяют три основные группы микроорганизмов.
1.Психрофилы – растут при температурах ниже +20 0С.
2.Мезофиллы – растут в диапозоне температур от 20 до 45 0С (часто оптимум – при +370С).
3.Термофилы – растут при температурах выше + 45 0С.
Краткая характеристика питательных сред. По консистенции
выделяют жидкие, плотные (1,5-3%-ныйагар) и полужидкие (0,3- 0,7 % агар) среды.
Агар – полисахарид сложного состава из морских водорослей, основной отвердитель для плотных (твердых) сред. В качестве универсального источника углерода и азота применяют пептоны – продукты ферментации белков пепсином, различные гидролизаты (мясной, рыбный, казеиновый, дрожжевой и др.).
По назначению среды разделяют на ряд групп:
универсальные (простые), пригодные для различных нетребовательных микроорганизмов(мясо-пептонныйбульон (МПБ), мясопептонный агар (МПА);
специальные – среды для микроорганизмов, не растущих на универсальных средах (средаМак-Коядля выделения возбудителя туляремии, средаЛевенштейна-Йенсена– возбудителя туберкулеза);
дифференциально-диагностические– для дифференциации микроорганизмов по ферментативной активности и культуральным свойствам (среды Эндо, Плоскирева, Левина, Гисса);
селективные (элективные) – для выделения определенных видов микроорганизмов и подавления роста сопутствующих – пептонная вода, селенитовая среда, среда Мюллера.
По происхождению среды делят на естественные, полусинтетические и синтетические.
Характер роста бактерий на плотных и жидких питательных средах: сплошной рост, образование колоний, осадок, пленка, помутнение.
Чистая культура – популяция одного вида микроорганизмов. Основные принципы получения чистых культур: механическое
разобщение, рассев, серийные разведения, использование элективных сред, особых условий культивирования (с учетом устойчивости некоторых микробов к определенным температурам, кислотам, щелочам, парциальному давлению кислорода, рН и др).
Лекция 7
ОБЩАЯ ВИРУСОЛОГИЯ
Открытие вирусов Д.И. Ивановским в 1892 г. положило начало развитию науки вирусологии. Быстрому ее развитию способствовали изобретение электронного микроскопа, разработка метода культивирования микроорганизмов в культурах клеток.
Слово «вирус» в переводе с латинского – яд (животного происхождения). Этот термин применяют для обозначения уникальных представителей живой природы, не имеющих клеточного (эукариотического или прокариотического) строения и обладающих облигатным внутриклеточным паразитизмом, т.е. не могут жить без клетки.
В настоящее время вирусология – бурно развивающаяся наука, что связано с:
ведущей ролью вирусов в инфекционной патологии человека (примеры: вирус гриппа, ВИЧ – вирус иммунодефицита человека, цитомегаловирус и другие герпесвирусы) на фоне практически полного отсутствия средств специфической химиотерапии;
использованием вирусов для решения многих фундаментальных вопросов биологии и генетики.
Вирусы – доклеточная форма существования живой материи, облигатные внутриклеточные паразиты, способные только к внутриклеточному размножению. Вирусы относятся к царству Vira.
Таксономические категории. Семейство имеет окончание viridae,
подсемейство – virinae, род – virus. Названия родов подсемейств даны не для всех вирусов. Вид вируса не получил биноминального названия, как у бактерий.
В основу классификации вирусов положены категории:
Тип НК (ДНК или РНК), ее структура, количество нитей (1 или 2).
Особенности репликации вирусного генома. Размер и морфология вирионов.
Количество капсомеров и тип симметрии нуклеокапсида. Наличие оболочки (суперкапсида).
Место размножения в клетке. Антигенные свойства и др.
Основные свойства вирусов. Ультрамикроскопические размеры, нм (от 15-20 до 350-400 нм). Один тип нуклеиновой кислоты – ДНК или РНК. Отсутствие способности к росту и бинарному делению. Размножение за счет воспроизводства себя из собственного генома. Отсутствие собственных систем мобилизации энергии. Отсутствие способности к синтезу белка. Среда обитания: бактерии, клетки растений, животных и человека.
Структура вириона. Различают: просто устроенные вирусы
(вирусы полиомиелита, гепатита А), которые состоят из нуклеиновой кислоты и капсида; сложно устроенные вирусы (вирусы кори, гриппа, герпеса, коронавирусы), состоящие из нуклеиновой кислоты, капсида и липопротеиновой оболочки (суперкапсид). В центре вириона – нуклеиновая кислота, окруженная капсидом. Капсид состоит из белковых субъединиц – капсомеров. Количество капсидов и способ их укладки строго постоянны для каждого вида вируса (вирус полиомиелита – 32 капсомера, аденовирус – 252). Суперкапсид у сложных вирусов образуется при выходе его из клетки хозяина. Вирусный капсид обволакивается внутренней поверхностью цитоплазматической мембраны клетки хозяина, и образуется один или несколько слоев оболочки – суперкапсид (вирусы бешенства, герпеса, энцефалита). У некоторых вирусов от внешнего липидного слоя оболочки выступают капсомеры в виде шипов – пепломеры (вирус гриппа). Нуклеиновая кислота вируса является носителем наследст-
венных свойств. Капсид и суперкапсид несут защитные функции, а также способствуют проникновению вируса в клетку.
Морфология вириона. В зависимости от способа укладки капсида различают формы: кубическую (икосаэдрическая); сферическую; палочковидную; пулевидную; спермазоидную.
Химический состав вирусов. Количество и содержание ДНК и РНК вирусов не одинаковы. Белки содержатся в незначительном количестве, состоят из16-20-иаминокислот. Функции: антигенные свойства, защита от ферментов клетки хозяина. Липиды и углеводы содержатся во внешней оболочке вирионов. Их источником является оболочка клетки хозяина. Ферменты вирусов. У большинства вирусов нет ферментов, т.к. они не имеют собственного метаболизма. Некоторые вирусы имеют ферменты, способствующие их проникновению в клетку хозяина (вирус гриппа А – нейраминидаза, фаги – лизоцим).
Пути передачи вирусов: воздушно-капельный(грипп, оспа), алиментарный (полиомиелит, гепатит А),контактно-бытовой(бешенство), трансмиссивный (энцефалит).
Пути проникновения вируса в клетку хозяина: рецептор-
зависимый эндоцитоз (виропексис), слияние оболочки вируса с клеточной мембраной.
Рецептор-зависимый эндоцитоз (виропексис): сначала идет ад-
сорбция вириона и клетки и далее захватывание и поглощение вириона клеткой. Клеточная мембрана с прикрепленным вирионом впячивается, образуется внутриклеточная вакуоль с вирусом. Содержимое вакуоли закисляется, вирусный нуклеокапсид выходит в цитоплазму клетки-хозяина.
Слияние оболочки вируса с клеточной мембраной характерно только для сложных вирусов (белки слияния). Происходит точечное взаимодействие вирусного белка слияния с липидами клеточной мембраны, вирусная липопротеиновая оболочка интегрируется с клеточной мембраной, а внутренний компонент попадает в цитоплазму клетки-хозяина.
Репликация вируса в клетке хозяина:
«Раздевание вируса» (депротеинизация). Удаляются поверхностные структуры: оболочка и капсид, высвобождается вирусная НК и транспортируется к местам депротеинизации.
Синтез вирусных компонентов. Происходит репликация НК и синтез вирусных белков при участии ДНК или РНК клетки хозяина.
Формирование вирусов – самосборка белковых частиц вокруг вирусной НК.
У одних вирусов самосборка происходит в цитоплазме, у других – в ядре клетки хозяина.
Образование внешней оболочки всегда происходит в цитоплазме.
Выход вирусов из клетки хозяина:
Взрывной путь: из погибающей клетки одновременно выходит большое количество вирионов. По взрывному пути выходят из клетки вирусы, не имеющие липопротеиновой оболочки.
Почкование, экзоцитоз характерно для сложных вирусов. Образовавшийся нуклеокапсид вириона транспортируется к мембранам клетки, в области контакта нуклеокапсида вириона с клеточной мембраной начинается выпячивание этих участков. Клетка способна длительно сохранять жизнеспособность и продуцировать вирусное потомство.
Типы взаимодействия вируса и клетки хозяина:
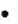 Продуктивный тип завершается образованием нового поколения вирионов и гибелью зараженных клеток. Гибель (лизис) клеток (цитопатический эффект) – результат интенсивного размножения и формирования большого количества вирусных частиц. Это характерный результат продуктивного процесса, вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит достаточно узнаваемый специфический характер; стабильное взаимодействие, не приводящее к гибели клетки (персистирующие и латентные ин-
Продуктивный тип завершается образованием нового поколения вирионов и гибелью зараженных клеток. Гибель (лизис) клеток (цитопатический эффект) – результат интенсивного размножения и формирования большого количества вирусных частиц. Это характерный результат продуктивного процесса, вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит достаточно узнаваемый специфический характер; стабильное взаимодействие, не приводящее к гибели клетки (персистирующие и латентные ин-
фекции) – так называемая вирусная трансформация клетки.
Абортивный тип – новые вирионы не образуются, т.к. инфекционный процесс прерывается на одном из этапов. Когда клетки освобождаются от вируса при инфицировании дефектным вирусом, для репликации которого нуженвирус-помощник,самостоятельная репликация этих вирусов не возможна. Например, вирус дельта (D) гепатита может реплицироваться только при наличии вируса гепатита B, егоHbs-антигена,аденоассоциированный вирус – в присутствии аденовируса); при инфицировании вирусом генетически не чувствительных к нему клеток; при заражении чувствительных клеток вирусом в определенных условиях.
Интегративный тип, или вирогения – встраивается вирусная ДНК в хромосому клетки и совместно сосуществует. Обеспечивается синхронность репликации вирусной и клеточной ДНК. Встраиваться в ДНК-геном хозяина могут только ДНК-вирусы (принцип «ДНК в ДНК»). Единственные РНК-вирусы, способные интегрироваться в геном клетки хозяина, – ретровирусы – имеют для этого специальный механизм. Особенность их репродукции: синтез ДНК провируса на основе геномной РНК с помощью фермента обратной транскриптазы с последующим встраиванием ДНК в геном хозяина.
Генетика вирусов. Геном вирусов содержит от 6 до нескольких сотен генов. Представлен различными видами нуклеиновых кислот:2-нитевыми,1-нитевыми,линейными, кольцевыми, фрагментированными. СредиРНК-содержашихвирусов различают:
Вирусы сплюс-нитьюРНК: (+)нить несет наследственную информацию и функцию иРНК.
Вирусы с(-)нитьюРНК:(-)нитьвыполняет только наследственную функцию.
Мутации вирусов:
Мутации могут возникнуть во время репликации нуклеиновой кислоты. Природа мутагенов: физические и химические факторы.
Генетическая рекомбинация – одновременное заражение клетки хозяина двумя вирусами, при этом возможен обмен отдельными генами между вирусами. Образуются рекомбинанты, содержащие гены двух родителей.
Генетическая реактивация генов происходит при скрещивании инактивированного вируса с полноценным, что приводит к спасению инактивированного вируса.
Методы культивирования вирусов:
вирусы размножаются только в жизнеспособных клетках; вирусы культивируют в куриных эмбрионах;
вкультурах ткани человека и животных;
ворганизме чувствительных животных.
Культивирование вирусов в куриных эмбрионах проводится в разных местах эмбриона: на хорион-аллантоисную оболочку; в аллантоисную полость; амниотическую полость; желточный мешок. Температурный режим и длительность инкубации зависит от биологических свойств введенного вируса.
Культивирование вирусов в культуре клеток
Используют ткани человека и различных животных. Однослойные культуры клеток выращивают в стеклянных плоских сосудах-матрицах. Присутствие вирусов в культурах тканей обнаруживают по изменению клеток. Тип вируса определяют реакцией нейтрализации типоспецифическими сыворотками.
Типы клеточных культур:
1. Первичные (трипсинизированные) культуры – фибробласты эмбриона курицы (ФЭК), человека (ФЭЧ), клетки почки различных животных и т.д. Первичные культуры получают из клеток различных тканей чаще путем их размельчения и трипсинизации. Используют их однократно, т.е. постоянно необходимо иметь соответствующие органы или ткани.
2. Линии диплоидных клеток пригодны к повторному диспергированию и росту, как правило, не более20-ипассажей (т.к. теряют исходные свойства).
3. Перевиваемые линии (гетероплоидные культуры), способны к многократному диспергированию и перевиванию, т.е. к многократным пассажам, наиболее удобны в вирусологической работе, например, линии опухолевых клеток Hela, Hep и др.
Специальные питательные среды для культур клеток
Используются разнообразные синтетические вирусологические питательные среды сложного состава, включающие большой набор различных факторов роста – среда 199, игла, раствор Хэнкса, гидролизат лактальбумина. В среды добавляют стабилизаторы рН (Hepes), различные в видовом отношении сыворотки крови (наиболее эффективной считают эмбриональную телячью сыворотку), L- цистеин иL-глютамин.
В зависимости от функционального использования среды могут быть ростовые (с большим содержанием сыворотки крови), их используют для выращивания клеточных культур до внесения вирусных проб, и поддерживающие (с меньшим содержанием сыворотки или ее отсутствием) для содержания инфицированных вирусом клеточных культур.
Проявления вирусной инфекции клеточных культур:
1.Цитопатический эффект.
2.Выявление телец включений.
3.Выявление вирусов методом флюоресцирующих антител (МФА), электронной микроскопией, авторадиографией.
4.Цветная проба. Обычный цвет используемых культуральных сред, содержащих в качестве индикатора рН феноловый красный, при оптимальных для клеток условиях культивирования (рН около 7,2) – красный. Размножение клеток меняет рН и, соответственно, цвет среды с красного на желтый за счет смещения рН в кислую сторону. При размножении в клеточных культурах вирусов происходит лизис клеток, изменения рН и цвета среды не происходит.
5.Выявление гемагглютинина вирусов – гемадсорбция, гемагглютинация.
6.Метод бляшек (бляшкообразование). В результате цитолитического действия многих вирусов на клеточные культуры образуются зоны массовой гибели клеток. Выявляют бляшки – вирусные «клеточно-негативные»колонии.
Методы идентификации вирусов: РГА (реакция гемагглютина-
ции); РТГА (реакция торможения гемагглютинации); РНГА (реакция непрямой гемагглютинации); РПГА (реакция прямой гемагглютинации); РСК (реакция связывания комплемента).
Бактериофаги
Бактериофаги – вирусы бактерий. Фаги могут инфицировать и разрушать только молодые развивающиеся клетки, являясь их паразитами.
Морфология фагов: фаги состоят из головки и хвостового отростка. Размеры фагов, форма и величина головки, длина и строение отростка различны у разных фагов.
Химический состав фагов. Как и все вирусы, фаги состоят из НК одного типа и белка. Молекула НК, скрученная в спираль, находится в головке фага. Капсид фага и отросток имеют белковую природу. На свободном конце отростка содержится литический фермент (лизоцим).
Специфичность фагов. Фаги обладают строгой специфичностью. Различают виды специфичности: видовая – способны паразитировать только в определенном виде бактерий (стрептококковый, стафилококковый, холерный и т.д.); типовая – способны паразитировать только на определенных представителях данного вида; поливалентная – способность лизировать бактерии близких видов (виды из рода возбудителей дизентерии (шигелл).
Взаимодействие фага с клеткой. Весь цикл взаимодействия фага с клеткой проходит в разных системах «фаг-бактерия» от нескольких минут до 1-2 ч в несколько стадий:
адсорбция – с помощью нитей хвостового отростка фаг адсорбируется на поверхностных рецепторах клетки. Адсорбция фагов специфична;
проникновение – стержень фага «прокалывает» клеточную стенку. Фермент, находящийся в отростке (лизоцим), разрушает ЦПМ. Чехол отростка сокращается, через канал стержня НК фага «впрыскивается» в клетку. Пустая белковая оболочка остается снаружи;
репродукция – репродукция белка и НК фага происходит внутри клетки;
сборка и формирование зрелых частиц фага;
лизис клетки и выход зрелых частиц фага из нее. Разрывается клеточная стенка, и в окружающую среду выходит несколько сотен новых фагов, способных поражать свежие клетки.
Практическое применение фагов: фагопрофилактика и фаготе-
рапия – предупреждение и лечение инфекций с помощью фагов. Фагодиагностика: идентификация выделенных культур; определение неизвестного фага по тест-культуремикробов; ускоренный метод диагностики инфекций с помощью реакции нарастания титра фага.
Лекция 8








