Вскоре после переоткрытия законов Менделя на животных и растениях разных видов было установлено, что не все признаки проявляют полное доминирование. Были выявлены случаи промежуточного наследования, неполного доминирования, сверхдоминирования и кодоминирования.
Если при скрещивании родительских особей с альтернативными признаками в потомстве появляются промежуточные фенотипы, то такое наследование называется частичным или неполным доминированием. Расщепление по фенотипу в F2 совпадает с расщеплением по генотипу, а именно 1:2:1.
Кодоминирование - Совместная экспрессия двух аллелей одного гена, кодирующих два различных продукта в гетерозиготном генотипе. Пример – наследование антигенных групп человека А, В, АВ и 0, детерминируемых геном I.
Пример наследования групп крови иллюстрирует и явление множественного аллелизма: ген I представлен тремя разными аллелями, кот комбинируются в зиготах только попарно.
Летальные гены. Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот F2. Разная жизнеспособность зигот может быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству. Летальные гены могут быть доминантными и рецессивными, но в большинстве случаев рецессивны, и поэтому долгое время могут находиться в скрытом состоянии. Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании.
Теория сверхдоминирования связывает гибридную мощность с преимуществом гетерозисного состояния (АА <Аа >аа). Эффект сверхдоинирования в гетерозиготе может наблюдаться даже в том случае, когда рецессивная аллель в гомозиготе летальна/приводит к снижению жизнеспособности.
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом поколении появляется новый признак, которого не было у исходных родительских форм, и соотношение фенотипов во втором поколении будет иным.
Комплементарное взаимодействие генов. В том случае, когда признак образуется при наличии двух доминантных неаллельных генов, каждый из которых не имеет самостоятельного фенотипического выражения, гены обозначают как комплементарные.
Эпистаз - Явление подавления действия одного гена другим, неаллельным ему геном. Виды: доминантный (A>B, A>b; 12:3:1) и рецессивный (a>B; a>b; 9:3:4). Ген, подавляющий действие другого, называют эпистатическим, ингибитором или супрессором. Подавляемый ген носит название гипостатического.
Полимерия. При полимерии, или полимерном (полигенном) наследовании, на один и тот же признак влияют несколько разных, но сходно действующих неаллельных генов. Каждый из них усиливает развитие признака. Такие однозначно действующие гены называются аддитивными.
Существует кумулятивная полимерия (степень проявления признака зависит от суммарного действия нескольких генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак). По такому типу наследуются количественные признаки: цвет кожи, молочность, яйценоскость, масса, длинна колоса и т.д. Расщепление в F2 по фенотипу при дигибридном скрещивании происходит в соотношении 1:4:6:4:1.
Некумулятивная полимерия: При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление в F2 по фенотипу при дигибридном скрещивании — 15:1.
В основе взаимодействия неаллельных генов лежит биохимическое отношение между белками-ферментами, которые кодируются комплементарными или эпистатическими генами.
У белков есть несколько функциональных центров: для связывания субстрата, для взаимодействия с кофакторами, с регуляторными молекулами, мембранами клетки и т.д. Каждый функц.центром представлен отдельным доменом – полуавтономным участком специфически уложенной полипептидной цепи. Разные рецессивные аллели одного и того же гена могут отлчаться друг от друга тем, что кодируют полипептиды с повреждениями различных доменов. Если аллели с разными повреждениями объединятся в гетерозготе т.о., что в молекуле фермента, состоящей из субъединиц-продуктов одного и того же гена, соберутся все необходимые функц.центры, то ферментативная активность будет восстановлена.
На примере дрозофил: окраска глаз обусловлена синтезом двух пигментов – красного и бурого. Рецессивная аллель bw (коричн цвет глаз) в гомозиготе прерывает синтез красного пигмента, поэтому глаза содержат только коричневый пигмент. И наоборот, рецессивная аллель st прерывает синтез бурого пигмента => глаза красные. Когда в дигетерозиготе оказываются нормальные аллели обоих генов, синтезируются оба пигмента. Результат – комплементарное взаимодействие нормальных аллелей. Если в гомозиготе оказываются и bw bw и st st, то не синтезируются ни красный, ни бурый пигменты, глаза белые.
Само словосочетание взаимодействие генов условно – взаимодействуют не сами гены, а их продукты.
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ № 3
Основы гибридологического метода: выбор объекта, отбор материала для скрещиваний, анализ признаков, применение статистического метода. Разрешающая способность гибридологического метода. Генетическая символика.
Метод был основан Г. Менделем в 1865г. Он положил в основу изучения наследования новый принцип: анализ наследования отдельных пар признаков в потомстве скрещиваемых растений 1 вида (горох), отличающихся по 1, 2 и 3 парам контрастных, альтернативных признаков (в отличие от его предшественников, которые изучали совокупности всех признаков у гибридного потомства).
Особенности:
1. Скрещиваемые организмы должны принадлежать к одному виду.
2. Скрещиваемые организмы должны четко разделяться по отдельным признакам.
3. Изучаемые признаки должны быть константны, т.е. воспроизводиться из поколения в поколение при скрещивании в пределах линии (родительской формы).
4. Необходимы характеристика и количественный учет всех классов расщепления, если они наблюдается у гибридов первого и последующих поколений.
Статистический метод:
Для оценки отклонения от теоретически ожидаемого результата используют метод 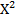 (хи-квадрат): =∑
(хи-квадрат): =∑ 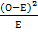 , где О – наблюдаемое, Е – ожидаемое. После расчета, необходимо оценить значимость отклонения по степени свободы: число классов (k) – 1, если значение не превышает табличного, то исходная гипотеза подтверждается.
, где О – наблюдаемое, Е – ожидаемое. После расчета, необходимо оценить значимость отклонения по степени свободы: число классов (k) – 1, если значение не превышает табличного, то исходная гипотеза подтверждается.
Разрешающая способность (эффективность) гибридологического метода определяется максимально возможным числом потомков, которые могут быть исследованы в анализирующих скрещиваниях. Поэтому первоначальные успехи в генетическом картировании эукариот были достигнуты при изучении дрозофилы, кукурузы, мыши и некоторых других видов организмов, характеризующихся относительно высокой разрешающей способностью анализа. Классический генетический анализ практически невозможен для организмов, экспериментальное размножение которых в достаточных количествах связано со значительными техническими трудностями. Гибридологический метод не применим также к человеку, за исключением возможности проводить гибридизацию его культивируемых соматических клеток.
Генетическая символика:
Скрещивание – знак умножения (×)
На первом месте женский пол - ♀, затем мужской - ♂
Родительские организмы - Р
Потомство от скрещивания 2х особей с различными признаками – гибридное, отдельная особь – гибрид. Гибридное поколение – F с индексом, соответствующим порядковому номеру поколения.
A — доминантный ген, а — рецессивный ген; АА — гомозигота по доминанте, аа — гомозигота по рецессиву, Аа — гетерозигота.
2. Закономерности нехромосомного наследования, отличие от хромосомного наследования. Методы изучения: реципрокные, возвратные и поглощающие скрещивания, метод трансплантации, биохимические методы.
Нехромосомное наследование – передача в ряду поколений генов, локализованных вне ядра. Для него характерны сложные картины расщепления, не согласующиеся с законами Менделя. Часто его называют цитоплазматическим наследованием (наследуемые гены расположены в цитоплазме, пластидах, митохондриях, инородных генетических элементах – вирусах)
!!! отличать от собственно цитоплазматического наследования (только цитоплазма несет гены)
Закономерности:
1. Частое соматическое (митотическое) расщепление;
2. Отсутствие или нерегулярное расщепление в мейозе;
3. У многих видов животных и растений имеет место однородительская передача признаков, определяемых структурами цитоплазмы.
Отличия от хромосомного наследования:
1. различия в результатах реципрокного скрещивания;
2. наличие связи между наследованием определенных признаков и переносом в клетку определенной цитоплазматической ДНК;
3. невозможность выявить сцепленность определенных генов с хромосомными генами;
4. отсутствие типичного количественного расщепления признаков в потомстве в соответствии с законами Менделя, зависимого от расхождения гомологичных хромосом в мейозе;
5. независимость проявления признака от замены ядер в клетках.
Реципрокное скрещивание, система из двух скрещиваний — прямого и обратного. При Р. с. каждый из генотипически различных родительских типов А и В используется дважды — один раз в качестве материнской и другой раз в качестве отцовской форм (♀А ´♂В и ♀В ´♂А). Различия между реципрокными гибридами могут быть вызваны влиянием материнского организма, цитоплазматической наследственности, сцепленными с полом генами.
Метод возвратного скрещивания состоит в получении потомства в ряду поколений от скрещивания гетерозиготы (детей гомозиготных родителей, генетически отличающихся друг от друга) с одним из исходных гомозиготных родителей. Смысл подобного скрещивания - замена гена (генов какого-либо комплекса) одной инбредной линии на гаплотип другой. В результате получается конгенная линия, отличающаяся от исходной только по этому гену (генам этого комплекса).
Поглотительное скрещивание, преобразовательное скрещивание, один из видов скрещивания, применяемый для коренного улучшения малопродуктивных пород высокопродуктивными. Простое П. с. заключается в спаривании животных двух пород (улучшаемой и улучшающей) для получения помесей, которых затем в ряде поколений спаривают с производителями улучшающей породы до получения животных желательного типа. Высокопродуктивных помесей 4—5—6-го поколений (высококровных), отвечающих типу улучшающей породы, разводят «в себе» (см. Разведение «в себе»), что иногда заканчивается созданием новой породы. П. с., в котором участвуют несколько улучшающих пород, называемых сложным. П. с. — наиболее быстрый и эффективный способ массового улучшения малопродуктивного скота, а также преобразования пород с.-х. животных (например, грубошёрстных пород овец в тонкорунных и полутонкорунных). Скорость преобразования и улучшения пород зависит от степени наследственных различий между животными скрещиваемых пород, степени наследственной устойчивости (консолидации) пород, тщательности отбора и подбора среди помесей, а также условий кормления и содержания помесного молодняка.
Методы трансплантации ядер: был разработан метод менее травматического переноса ядер методом микроманипуляции. Он протекает в два этапа: сначала тонкой микропипеткой прокалывают зоны пеллюцида и плазматической мембраны и извлекают пронуклеусы, а затем другой пипеткой, большего диаметра (12 мкм) в то же отверстие вводят диплоидное ядро донора. В этом случае меньше травмируется цитоплазма зиготы и транспортируемое ядро донора. Трансплантация ядер может осуществляться и другим способом, с использованием цитохалазинов (веществ, синтезируемых грибами). Цитохалазин В разрушает структуру микрофиламентов и способствует уникальному расположению ядра. Ядро остается соединенным с клеткой тоненьким стебельком цитоплазмы. При центрифугировании этот мостик разрывается, образуются безъядерные клетки (цитопласты) и кариопласты, представляющие собой ядра, окруженные тонким слоем цитоплазмы и цитоплазматической мембраной. Цитопласты отделяют от интактных клеток в градиенте плотности. Они сохраняют способность прикрепляться к поверхности культурального сосуда и могут быть использованы для слияния с кариопластами других клеток с целью получения жизнеспособной клетки.
Биохимические методы направлены на выявление биохимического фенотипа организма. Биохимические показатели (первичный белковый продукт гена, накопление патологических метаболитов внутри клетки) отражают сущность болезни более адекватно, чем клинические симптомы. С помощью биохимических методов описано более 1000 врожденных болезней обмена веществ.
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ № 4
Хромосомные перестройки. Внутри- и межхромосомные перестройки; делеции, дупликации, инверсии, транслокации, транспозиции. Механизмы их возникновения, использование в генетическом анализе для локализации отдельных генов и составления генетических карт. Особенности мейоза при различных типах перестроек.
Хромосомные мутаци, или хромосомные перестройки, или хромосомные аберрации представляют собой перемещение ген. материала, приводящие к изменению структуры хромосом в пределах кариотипа. Выделяют аберрации внутрихромосомные и межхромосомные.
Внутрихромосомные: дефишенси, делеции, дупликации, инверсии, транспозиции*.
Межхромосомнве: транслокации, транспозиции*.
*Транспозиции занимают промежуточное положение м/у внутри- и межхр. перестройками, т.к. они могут происходить как м/у негомологичными хр-ми, так и в пределах одной хр.
1)Делеции – выпадения частей хромосомы, не захватывающие теломеру, а также концевые нехватки, первоначально обозначавшиеся термином дефишенси, который теперь используют редко.
Вследствие нехваток хромосомы укорачиваются, и физическое отсутствие участка одного из гомологов приводит к гемизиготному состоянию генов, находящихся в нормальном гомологе. Если теряются доминантные аллели одного из гомологов гетерозиготы, то наблюдают фенотипическое проявление рецессивных аллелей хромосомы, не затронутой аберрацией. Например, у дрозофилы в Х-хромосоме известна доминантная мутация (делеция локуса W) Notch, приводящая в гетерозиготе к появлению вырезки на крыле. В гомозиготном или гемизиготном состоянии она летальна.
Поскольку вследствие делеций теряются участки хромосом, у гетерозигот по этим перестройкам наблюдаются характерные нарушения конъюгации гомологов. Более длинная нормальная хромосома образует петлю на участке, соответствующем делеции. Это часто используют при построении генетических карт.
Концевые нехватки, или дефишенси, устанавливают по тем же критериям, что и делеции, однако вследствие их расположения при конъюгации не образуется петля, а одна хромосома оказывается короче другой. Пример у человека – синдром кошачьего крика.
Большие возможности для выявления делеций, дефишенси и других хромосомных аберраций открывает метод дифференциальной окраски хромосом. Благодаря этому хромосомы приобретают характерную поперечную исчерченность.
2)Дупликация – удвоение (умножение) части хромосомы. Известны случаи многократных повторений, или мультипликаций, какого-либо участка. Их называют также амплификациями. Дупликации могут происходить в пределах одной и той же хромосомы или сопровождаться переносом копии участка генетического материала на другую хромосому. Повторы, возникшие в одной хромосоме, могут располагаться тандемно (ABC BC DE…) или инвертированно (ABC CB DE…). Различают также терминальные повторы, если дупликация затрагивает конец хромосомы.
По-видимому, главной причиной множественных повторов участков генетического материала является неравный кроссинговер (локус Bar у дрозофилы).
Фенотипическое проявление дупликаций показано на примере ряда генов у дрозофилы, как рецессивных: white (w), scute-achaete (sc-ac) – редукция щетинок, vermilion (v) – ярко-красные глаза, так и доминантных: Bar (B), Hairy wing (Hw) – волосатые крылья.
Гетерозиготы по дупликациям выявляют сходно с гетерозиготами по делециям – образуется петля при конъюгации хромосом.
На существование повторов в геномах других организмов указывает, например, спаривание отдельных участков негомологичных хромосом в мейозе у гаплоидных растений, которые могут быть получены экспериментальным путем. В отсутствие гомологов у гаплоидов наблюдают участки синапсиса – гомологическое спаривание.
При дупликации доминантной аллели какого-либо гена, сопровождаемой транспозицией копии на другую хромосому, меняется и картина расщепления, если скрещивание проводят с рецессивным мутантом по тому же гену. Вместо моногибридного расщепления 3:1 у диплоидных организмов наблюдают полимерное расщепление 15:1.
3)Инверсия – изменение чередования генов в хромосоме вследствие поворота участка хромосомы на 180°. Этот тип перестроек чаще всего встречается в природных популяциях. В зависимости от расположения концов перестройки по отношению к центромере инверсии делят на перицентрические, включающие центромеру в инвертированный участок, и парацентрические, не включающие центромеру в инвертированный участок.
Инверсия приводит к изменению сцепления генов, иной их линейной последовательности, нежели у исходной формы. Этот эффект можно обнаружить, если инверсия в гомозиготе не летальна. Рецессивная летальность часто сопутствует инверсиям как результат локализации точек разрывов в жизненно важных генах или как следствие эффекта положения.
Другое важное следствие инверсии – подавление кроссинговера, если инверсия находится в гетерозиготе. У гетерозигот по инверсиям на цитологических препаратах обнаруживают характерные петли – результат конъюгации структурно измененной и нормальной хромосомы. Двойной кроссинговер у гетерозигот по инверсии может приводить к образованию вполне жизнеспособных гамет.
Хромосома может нести не только одну инверсию, но две неперекрывающиеся и две, перекрывающиеся полностью или частично. Гетерозиготность по таким сложным перестройкам также идентифицируют цитологически по характеру конъюгации хромосом.
4)Транслокации – реципрокный (взаимный) обмен участками негомологичных хромосом. В результате такого обмена у гомозигот по транслокациям изменяется характер сцепления генов. В гетерозиготе по транслокации гены, принадлежавшие разным, негомологичным хромосомам, наследуются как принадлежащие одной группе сцепления. Это объясняется тем, что полностью функциональными оказываются только те споры и гаметы, которые несут родительские сочетания хромосом.
Характер конъюгации транслоцированных хромосом меняется: образуется фигура креста. Плотная конъюгация вблизи точек разрывов оказывается затрудненной, что приводит к подавлению кроссинговера в этих участках.
У гетерозиготы по транслокации в профазе мейоза образуются квадриленты, а не биваленты, как обычно, поскольку гомологичные участки оказываются у всех четырех конъюгирующих хромосом. Из шести возможных типов гаплоидных продуктов, возникающих при трех способах расхождения хромосом, только два типа функционируют нормально – те, которые получают полные наборы генов, характерные для исходных родительских форм.
Остальные четыре типа несут дупликации и нехватки и потому, как правило, не дают жизнеспособного потомства или не участвуют в оплодотворении.
Характерный пример в этом отношении представляют разные виды растений ослинника.
5)Транспозиции – перемещение небольших участков ген. материала в пределах одной хр. или м/у разными хр-ми. Транспозиции происходят при помощи мигрирующих генетических элементов (МГЭ). МГЭ – это структуры, которые могут перемещаться в пределах генома и переходить из генома в геном. Они могут встраиваться в различные области хромосом эукариот, ДНК и плазмид прокариот. МГЭ были открыты Б. Мак-Клинток у кукурузы еще в 1940-е гг.
У прокариот выделяют два вида МГЭ:
Инсерционные последовательности (IS) – небольшие последовательности ДНК (700–2000 п. н.), имеющие на концах короткие (10–40 п. н.) схожие последовательности, расположенные в обратном порядке (инвертированные повторы).
Все IS-элементы характеризуются след. структурными особенностями:
1) На концах имеются инвертированные повторы;
2) Большинство содержит ген фермента транспозазы, ответственной за их перемещение;
3) IS-эл. могут содержать по неск. сигналов начала и конца трансляции, а также сочетание нуклеотидов, сходное с сигналами терминации транскрипции.
4) В точке внедрения каждого IS-эл., на его флангах всегда обнаруживается дупликация (в прямой ориентации), не являющаяся частью IS-эл. Представляет собой результат повторений сайта-мишени, в кот. внедряется.

Рис. Схема внедрения IS в ДНК хозяина:1 – удвоенный сайт-мишень ДНК хозяина; 2 – инвертированные повторы; 3 – центральная область IS; 4 – ДНК клетки-хозяина.
Бактериальная клетка обычно имеет множество IS. Наличие идентичных IS в различных бактериальных генетических структурах создает основу для их взаимодействия, интеграции и генетического обмена по принципу сайт-специфической рекомбинации.
Позже у бактерий были обнаружены более сложные мигририрующие элементы – транспозоны (Tn), которые отличаются от IS-эл. тем, что в них включены некоторые гены, не имеющие отношение к самому процессу транспозиции.
Вероятно, Tn возникли путем объединения двух первоначально независимых IS и области между ними (Shapiro J., 1983). Многие транспозоны несут гены устойчивости к антибиотикам. Процесс транспозиции у бактерий включает дупликацию транспозона, при этом одна копия остается на старом месте, а другая возникает на новом.
Обычно перемещение МГЭ не направлены и довольно редки. Они происходят с частотой около 1*10-5 – 1*10-8.
Предмет и методология селекции. Генетика как теоретическая ос нова селекции. Учение об исходном материале. Центры происхождения культурных растений по Н.И. Вавилову. Понятие о породе, сорте, штамме. Сохранение генофонда ценных культурных и диких форм растений и животных.
Селекция — это наука о путях создания новых и улучшения уже существующих сортов культурных растений, пород домашних животных и штаммов микроорганизмов с ценными для практики признаками и свойствами.
Теоретической базой этой науки является генетика. Селекция опирается на достижения молекулярной биологии, биохимии и др. наук о раст., жив. и мо. В последние годы селекция обогатилась результатами генной и клеточной инженерии, биотехнологии. Породы жив., сорта рас.т, штаммы мо предст-т собой совокупности особей, созданных чел с помощью методов селекции и харак-ся определ-ми наследственными особенностями, морфологическимии физиологическими хозяйственно ценными качествами. Поскольку свой-ва живых организмов обусловлены их нормой реакции на основе определенной генетической информации и подвержены модификационной и наследственной изменч-ти, развитие селекции основано на закономерностях генетики. Основные методы селекции вкл. отбор, гибридизацию, полиплоидию, мутагенез, клеточная и генная инженерия.
Очевидно, что для того, чтобы селекция была успешной, селекционер должен иметь коллекцию разных сортов и даже образцов дикорастущих форм того или иного вида, другими словами, обладать обширным генофондом видов растений, животных и микроорганизмов, селекцией которых он занимается.
Коллекцию разных сортов, форм и диких видов растений, от которых они произошли, называют генетическим банком. Созданием таких коллекций растений стали заниматься ещё на заре развития генетики. Основоположником таких исследований был Н. И. Вавилов. Вавилов в результате многичисленных экспедиций изучил многообразие и географическое распространение культурных растений, он установил, что для разных культур существуют свои центры многообразия, где сосредоточено наибольшее число сортов, разновидностей. Он выделяет 8 осн-х центров происхождения:1 индийский (рис, цитрус, сах тростник); 2 средиазиатский (горох, бобы, пшеница); 3 китайский (хлеб, злаки, бобовые); 4 средиземноморский (морская капуста, клевер); 5 абиссинский (кофе, ячмень); 6 переднеазиатский (пшеница, рожь, дыня); 7 южно-мексикансикй (хлопчатник, кукуруза, томаты, тыква); 8 южно-американский (картофель, хинное дерево, ананас). Центры происхождения важнейших культурных растений связаны с древними очагами цивилизации и местами первичного возделывания и селекции растений. Подобные очаги одомашнивания (центры происхождения) выявлены и у домашних животных.
Породой жив. или сортом раст. наз-т совокупность особей (популяцию), искусственно созданную чел, кот-ая характеризуется определенными наследственными особенностями: продуктивностью, морфологическими и физиологическими признаками. Штамм – чистая культура микроорг одного вида, у кот изучены морфол и физиол особенности.
Обитающие в каждом регионе виды животных, растений и микроорганизмов образуют целостную систему, известную как экосистема. Каждый вид представлен в ней своей, уникальной популяцией. Оценить экологическое благополучие данной территории или акватории позволяют данные, характеризующие генофонд ее экосистемы, т.е. генофонд слагающих ее популяций. Именно он обеспечивает существование экосистемы в данных условиях. Поэтому за изменениями в экологической обстановке региона можно проследить, изучая генофонды популяций обитающих там видов.
Осваивая новые территории, прокладывая нефте- и газопроводы, следует заботиться о сохранении и восстановлении природных популяций. Популяционная генетика уже предложила свои меры, например выделение природных генетических резерватов. Они должны быть достаточно обширными, чтобы содержать основной генофонд растений и животных данного региона. Теоретический аппарат популяционный генетики позволяет определить ту минимальную численность, которая необходима для поддержания генетического состава популяции, чтобы в ней не было т.н. инбридинговой депрессии, чтобы она содержала основные генотипы, присущие данной популяции, и могла воспроизводить эти генотипы. При этом каждый регион должен иметь свои собственные природные генетические резерваты. Нельзя восстанавливать загубленные сосняки Севера Западной Сибири, завозя семена сосны из Алтая, Европы или Дальнего Востока: через десятки лет может оказаться, что «чужаки» генетически плохо приспособлены к местным условиям. Вот почему экологически грамотное промышленное освоение территории должно обязательно включать популяционные исследования региональных экосистем, позволяющие выявить их генетическое своеобразие.
Сказанное относится не только к растениям, но и к животным. Генофонд той или иной популяции рыб эволюционно приспособлен именно к тем условиям, в которых он обитал в течение многих поколений. Поэтому интродукция рыб из одного природного водоема в другой порой приводит к непредсказуемым последствиям. Например, попытки развести сахалинскую горбушу в Каспии оказались безуспешными, ее генофонд оказался не в состоянии «освоить» новое местообитание. Та же горбуша, интродуцированная в Белое море, покинула его и ушла в Норвегию, образовав там временные стада «русского лосося».
Не надо думать, что основными объектами заботы о природе должны быть только экономически ценные виды растений и животных, такие, как древесные породы, пушные звери или промысловые рыбы. Травянистые растения и мхи, мелкие млекопитающие и насекомые – их популяции и их генофонды наравне со всеми другими обеспечивают нормальную жизнь территории. То же относится к микроорганизмам – тысячи их видов населяют почву. Изучение почвенных микробов – задача не только микробиологов, но и популяционных генетиков.
Изменение генофонда популяций при грубых вмешательствах в природу выявляется не сразу. Могут пройти десятилетия, прежде чем станут очевидными последствия в виде исчезновения одних популяций, а за ними – других, связанных с первыми.
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ № 5
1. Плазмидное наследование. Свойства плазмид: трансмиссивность, несовместимость, детерминирование признаков устойчивости к антибиотикам и другим лекарственным препаратам, образование колицинов и др. Использование плазмид в генетических исследованиях.
Признание за ядром главенствующей роли в передаче наследственных свойств не исключает существования внеядерной наследственности, которая связана с органоидами клетки, способными к саморепродукции. Факторы наследственности, расположенные в клетках вне хромосом, получили название плазмид. Функция плазмид, как и генов, находящихся в хромосомах, связана с ДНК.
Такие цитоплазматические структуры, как пластиды, митохондрии и центриоли способны к авторепродукции. Именно с ними связана передача цитоплазматической наследственности. Проявление этой формы наследственности находится под контролем ядерной ДНК.
Плазмиды — дополнительные факторы наследственности бактерий и некоторых простейших, расположенные в клетках вне хромосом и представляющие собой кольцевые (замкнутые) или линейные молекулы ДНК. Плазмиды способны реплицироваться автономно, но при этом они эксплуатируют репликационную систему клетки хозяина. Большинство плазмид имеет специальные белки - инициаторы репликации. Эти белки начинают процесс репликации, который затем подхватывается и продолжается репликационной системой клетки. Некоторые плазмиды могут обратимо встраиваться в бактериальную хромосому и функционировать в виде единого репликона. Такие плазмиды называются интегративными или эписомами.
Свойства плазмид:
1)Способность к автономной репликации - молекулы плазмидной ДНК приобретают ее в том случае, если в составе их имеется сайт начала репликации – oriV и набор генов, необходимых для ее осуществления. Различают плазмиды со строгим и ослабленным контролем репликации.
Плазмиды со строгим контролемрепликации удваиваются синхронно с бактериальной хромосомой при участии ДНК-полимераза III. Таких плазмид в клетке может насчитываться от 1 до 3. Репликация плазмид с ослабленным контролемпроисходит с участием ДНК-полимеразы I. В каждой клетке содержится 40–50 копий таких плазмид, их называют мультикопийными.
2) Трансмиссивность– способность плазмид передаваться из клетки в клетку при конъюгации. В зависимости от этого плазмиды деляют на конъюгативные (трансмиссивные) и неконъюгативные (нетрансмиссивные).
Конъюгативные плазмиды передаваются из одной клетки в другую, имеют гены, ответственные за перенос (tra-гены). Большинство конъюгативных плазмид имеет ограниченный круг клеток-хозяев. Меньшее число плазмид передаются широкому кругу клеток-хозяев (это космополитные).
Неконъюгативные плазмиды – мелкие плазмиды,не содержат tra-генов, неспособны самостоятельно передаваться от одних клеток к другим. Неконъюгативные плазмиды могут быть перенесены в реципиентные клетки с помощью конъюгативных плазмид. Перенос неконъюгативных плазмид с помощью конъюгативных называется мобилизацией. Мобилизация может функционировать из-за наличия в плазмидах IS-элементов и транспозонов, которые обеспечивают образование коинтегратов. В реципиентных клетках коинтеграты распадаются на два автономных репликона.
3) Способность плазмид к интеграции в бактериальную хромосому.
4) Несовместимость. Родственные плазмиды не могут сосуществовать в одной клетке, поскольку они несовместимы.
5) Поверхностное исключение присуще конъюгативным плазмидам. Свойство означает, что если в клетке есть определенная плазмида, то другая плазмидная ДНК при конъюгации с трудом преодолевает клеточную стенку.
Cуществует также множество специфических функций, детерминируемых той или иной плазмидой. у бактерий наиболее изучены три главные группы: F-плазмиды (факторы фертильности) ответственны за половой процесс, R-плазмиды (факторы резистентности) обеспечивают устойчивость бактериальных клеток к действию антибиотиков (к стрептомицину, тетрациклину) и сульфаниламидным препаратам, и Col-плазмиды (колициногенные факторы) в которых локализованы гены синтеза колицинов (бактериоцинов) - токсичных белков.
R-плазмиды. Обусловленная плазмидами устойчивость бактерий к антибиотикам основана на разных механизмах, но чаще всего - на инактивации последних ферментами, кодируемыми плазмидами или на избирательном изменении проницаемости клеточной оболочки.
Среди плазмид., обеспечивающих устойчивость бактерий к антибиотикам, основную массу составляют так называемые факторы множеств, резистентности, несущие сразу несколько соответствующих детерминант. С помощью трансмиссибельных плазмид детерминанты резистентности легко могут распространяться между видами, способными к конъюгации. На такие плазмиды гены резистентности могут передаваться с помощью транспозонов.
F-плазмиды называют еще половым фактором или фактором фертильности (откуда и название) потому что наличие F-плазмиды превращает клетку в возможного донора наследственного материала при конъюгации (т.е. в «мужскую» клетку), соответственно клетка, лишенная F-плазмиды, является потенциальным реципиентом генетического материала при конъюгации («женской» клеткой). Реципиентная клетка, получив при конъюгации F-плазмиду, превращается тем самым из женской в мужскую (т.е. происходит «смена пола»).
Col-плазмиды контролируют синтез особого рода антибактериальных веществ белковой природы - бактериоцинов, способных вызывать гибель бактерий того же вида или близких видов. Бактериоцины обнаружены у кишечной палочки (колицины), холерных вибрионов (вибриоцины), стафилококков (стафилоцины). Известно более 200 различных бактериоцинов.
Перспектива использования плазмид в связи с их исключительной биологической пластичностью оценивается молекулярными биологами трезво и объективно. Интенсивно ведется изучение в области генной инженерии. Плазмиды в ней широко используются для переноса генетической информации от донора к реципиенту и генетических манипуляций. Для этого создаются искусственные плазмиды — векторы, состоящие из частей, взятых из разных генетических источников, а также из искусственно созданных фрагментов ДНК.
Трансформируя гибридные плазмиды в клетки, удается повысить дозу генов и, следовательно, количество ферментов, ответственных за биосинтез какого-либо вещества.
В связи с существованием R-плазмид активно развивается новое направление в практической бактериологии - поиск и создание веществ, избирательно подавляющих репликацию плазмид или экспрессию их генов.








