Химический состав вод океана претерпевал изменения за время преобразования земной коры, особенно в период до 2 млрд лет назад. Океан служит резервуаром, в котором перемешиваются вещества в непрекращающемся процессе превращения изверженных пород в осадочные материалы, привноса ионов реками и ювенильными водами, а также перехода ионов в осадки. В. И. Вернадский (1926) писал: "Мировой океан с происходящими в нем сложными процессами равновесия не является изолированным в земной коре. Его вещество находится в теснейшем обмене с атмосферой и сушей, и этот обмен имеет огромное значение не только для химии моря, но и для химии всей земной коры".
Нормальные пределы изменения солености мирового океана - 33,0 - 37,0 %о, масса солей составляет 4,8 · 1018 т. Соленость на поверхности
открытого океана изменяется (в %0) от 32,5 в северной части Тихого океана и 34,0 в антарктических широтах до 36,0 в центрах интенсивного испарения в средних широтах Индийского океана и 37,0 в Атлантике, а в Персидском заливе - до 50- 100. Если все соли, содержащиеся в растворе, извлечь и ровным слоем разместить на поверхности земного шара, то его толщина составит 12м. Общее количество солей в океане составляет 5 · 1016 т. Практически все элементы таблицы Менделеева имеются в океанической воде, встречаясь прежде всего в виде соединений: силикатов, фосфатов, нитратов, нитритов, аммиака и органически связанного азота. Преобладают хлориды и карбонаты: на долю ионов хлора приходится 55,0% всех солей, карбонатов - 30,6, сульфатов - 7,7, магнезии - 3,7, кальция - 1,2, поташа - 1,1, брома - 0,2, бора - 0,07%.
Кроме биогенных элементов (N, Р, Si) важную физиологическую роль в процессе развития фитопланктона играют также некоторые каталитические микроэлементы, например, Fe, Cu, Mg, Со и др., выступая в качестве катализаторов окислительно-восстановительных процессов, протекающих в живом веществе, оказывают значительное влияние на процессы фотосинтеза и обмена веществ. Недостаточность этих микроэлементов в морской воде является фактором, лимитирующим развитие фитопланктона.
Принос в океан речным стоком биогенных элементов и микроэлементов, а также вынос их из глубинных слоев при подъеме вод и вертикальном перемешивании далеко не всегда компенсируют их недостаточность. Соответствующие расчеты количества микроэлементов, необходимых для обеспечения первичной продукции фитопланктона Мирового океана, показали, что в биологический круговорот вовлекаются и полностью (или даже неоднократно) используются поставляемые реками Ва, Ni, PI, Ag, Cd, Zn, Cu, Si. Очевиден напряженный баланс этих микроэлементов в океане. Распределение их концентраций по глубине свидетельствует о многократном превышении концентрации фосфатов и кремневой кислоты в глубинной зоне по сравнению с поверхностным слоем. Уже на глубине 200 м это преобладание обычно бывает 2 -4-кратным, а на глубине 800- 1000 м - еще большим. Круговорот кремния, фосфора и связанного азота почти целиком определяется процессом извлечения их фитопланктоном. Поступление растворенных форм этих элементов с речным стоком незначительно (менее 1 %).
Создание первичной продукции жестко лимитировано исключительной напряженностью "фонда" биогенных элементов. Наиболее продуктивными являются воды приарктических районов океанов, где зимой происходит интенсивная конвекция, а также приантарктических, где наблюдается постоянный подъем глубинных вод. Если в тонком поверхностном слое морей и океанов производится
органическое вещество, то оно потребляется как здесь, так и в более глубоко лежащих слоях. Биомасса живых организмов после отмирания почти полностью минерализуется и частично переходит в донные отложения, подчас надолго выпадая из процессов обмена в биосфере. Некоторая и с каждым годом все возрастающая доля биомассы водных организмов изымается человеком, что уменьшает объем минеральных фондов биогенных элементов.
Водные организмы используют для своего развития и воспроизводства множество различных элементов, причем некоторые из них включаются в процессы жизнедеятельности в микродозах. 99,9993 % массы находящихся в воде соединений приходится на 11 ионов основного солевого состава: Сl-, Na+, Mg2+, SO
, Ca2+, К+, Br-, HCO
, Sr2+, F- и HSO
(табл. 8). Именно они требуются для синтеза живых тканей в значительных количествах.
Органическая жизнь существует в природных водах любой солености от слабоминерализованной воды северных рек и озер
Таблица 8
Концентрация и время пребывания элементов в морской воде
(поТ.Шопфу, 1982)
| Элемент
| Концентрация, мг/л
| Продолжительность, годы
| Элемент
| Концентрация, мг/л
| Продолжительность, годы
|
| Н
|
| -
| К
|
| 6·106
|
| Li
| 0,18
| 2,3
| Ca
|
| 8,5·105
|
| Be
| 6·10-6
| -
| Mn
| 0,0002
| 104
|
| В
| 4,5
| 107
| Fe
| 0,001
| 102
|
| С
|
| -
| Cu
| 4·105
| 104
|
| N
|
| -
| Br
|
| 108
|
| О
| 880 000
| -
| Ag
| 1·105
| 4·104
|
| F
| 1,3
| 5 ·105
| I
| 0,06
| 4·105
|
| Na
|
| 4,8· 107
| Ba
| 0,02
| 4·104
|
| Mg
| 1 290
| 107
| La
| 3·10-6
|
|
| Al
| 0,001
| 102
| Au
| 4 ·10-7
| 2·105
|
| Si
| 3,0
| 1,8· 104
| U
| 0,03
| 3·106
|
| P
| 0,07
| 1,8· 105
| Tl
| 1·10-5
| 2·102
|
| S
|
| -
| Ra
| 7· 10-4
| 1·106
|
| Cl
|
| 108
|
|
|
|
(<0,1 %о) до водоемов с самосадочной солью (=300 %о). По отношению к солености различаются в качественном и количественном отношении главные типы водных организмов: морские, пресноводные, солоноватоводные и организмы пересолоненных водоемов.
Н.М.Книппович (1938) принимает за границу между морскими и солоноватыми водами соленость 24,7 ‰, при которой температура замерзания воды и температура ее наибольшей плотности одинаковы и составляют -1,33°С. Некоторые исследователи считают, что реальность установленной исключительно по физическому принципу границы Книпповича подтверждается фаунистическими материалами. Однако С. А. Зернов (1949) принимает за верхний предел солоноватых вод соленость 16‰, так как черноморская фауна до этой солености сохраняет типичный морской облик.
Всех водных животных подразделяют на две большие группы: пойкилосмотические, внутренняя среда которых (кровь, лимфа, гемолимфа, целомическая жидкость) находится в осмотическом равновесии с внешней средой, и гомойосмотические, поддерживающие давление внутренней среды на уровне, отличном от осмотического давления внешней среды.
К пойкилосмотическим животным относится подавляющее большинство морских беспозвоночных и немногочисленные группы позвоночных. Осмотическое давление внутренней среды беспозвоночных обеспечивается прежде всего минеральными ионами; доля органических веществ очень мала. Напротив, у пойкилосмотических позвоночных, способных к жизни в условиях переменной солености (зеленая жаба, таиландская лягушка-крабоед), осмотическое давление внутренней среды наполовину или даже больше обеспечивается органическими веществами, в первую очередь мочевиной. Таким образом, создается осмотическое "противодавление", препятствующее движению воды через клеточную мембрану и тем самым сохраняющее относительное постоянство объема клетки.
В биологических процессах, протекающих в морской воде, заметную роль играют некоторые газы. Из азота, кислорода и диоксида углерода, растворенных в воде в наибольших количествах, особое значение для поддержания жизни в океане имеет кислород. Содержание газообразного азота во всей толще вод океана сохраняется на уровне полного насыщения.
Воды Мирового океана пополняются кислородом преимущественно за счет жизнедеятельности фитопланктона и притока из атмосферы. Из 223,6 млрд т кислорода, поступающего в океан за год, 220 млрд т (98,4%) создается в результате жизнедеятельности растений и 3,6 млрд т (1,6 %) поступает из атмосферы. Поверхностные водные массы оказываются наиболее обогащенными кислородом,
и именно они снабжают им более глубокие слои воды океана. Опускание холодных водных масс, течения, круговороты, сезонные и суточные вертикальные их перемешивания способствуют обогащению кислородом глубинных вод Мирового океана.
На глубине водные массы постепенно теряют растворенный в них кислород. Поэтому, как правило, содержание кислорода в пределах абиссальных глубин сильно понижено, а в некоторых впадинах (в Черном, Балтийском морях, Норвежских фиордах и др.) он полностью отсутствует, что ограничивает или даже исключает растительную и животную жизнь.
В некоторые годы дефицит кислорода наблюдается в северозападной части Индийского океана. В период зимнего муссона опресненные поверхностные воды Бенгальского залива и Андаманского моря выносятся Северо-Восточным течением в Аравийское море. Образуют устойчивый слой, почти полностью прекращающий вертикальную конвекцию и способствующий быстрому образованию кислородного дефицита в подстилающих его слоях, приводящему к заморам и гибели рыбы; иногда масса погибшей рыбы достигает нескольких миллионов тонн.
Итак, кислородный режим является важным фактором, определяющим биопродуктивность вод. По самым ориентировочным подсчетам, примерно 0,6 млн км2 площади шельфа и верхней части склона в Мировом океане из-за крайне неблагоприятного кислородного режима исключается из числа перспективных районов для промысла придонных рыб.
Температура морской воды зависит от географической широты и сильно меняется в пространстве и во времени. Диапазон изменения составляет от 2 °С в высоких широтах до 60 °С в изолированных мелководных заливах в тропиках, однако на обширных пространствах тропического пояса температура составляет примерно 25 °С в поверхностном слое. Виды тропических морских организмов обитают в очень узком интервале температуры. Сезонные потепления и похолодания ограничены примерно 100-метровой толщей (сезонный термоклин), затем следует слой до 2000 м, где температура постепенно уменьшается (постоянный термоклин). Ниже идет глубинная водная масса с температурой 0 -4 °С. В самых глубоких слоях (6000- 11000 м) колебания температуры составляют от 1,1 до 3,6° С.
Годовые колебания температуры редко превышают 6 - 7°С, а в полярных водах 2 "С. Вода быстро охлаждается с глубиной и даже в тропиках на дне Атлантического океана температура составляет О -0,5°С, а в Белом море глубже 50 м вода всегда охлаждена до 1,4°С. В океанах градиенты температуры в местах соприкосновения водных масс создают "барьеры", непреодолимые для фауны. По мнению палеогеографов, температура поверхности океана существенно не менялась на протяжении последних 800 млн лет.
Диапазон температуры, в котором происходит нерест и эмбриональное развитие многих морских животных, оказывается часто более узким и четко выраженным, чем диапазон температуры, в котором возможна нормальная жизнедеятельность взрослых животных.
Поскольку процесс фотосинтеза инициируется солнечной радиацией, количество световой энергии в определенной точке океана, безусловно, лимитирует первичную продукцию органического вещества. Освещенность поверхностных слоев моря зависит от географической широты места, сезона года, облачности, высоты солнца над горизонтом и количества влаги в атмосфере. Интенсивность света быстро уменьшается с глубиной. В среднем на глубине 10м для фотосинтеза доступно лишь 10 % световой энергии, поступившей на поверхность моря, а на глубине 100 м - только 1 %. Зона первичного продуцирования ограничивается несколькими десятками метров от поверхности. В прибрежных водах, где обычно содержится значительно больше взвешенных частиц, чем в водах открытого океана, света на глубину проникает еще меньше.
Вода поглощает в большем количестве красные лучи спектра и относительно лучше пропускает сине-фиолетовые. В связи с этим зеленые и бурые водоросли, обитающие ближе к поверхности моря, сменяются на больших глубинах красными водорослями, окраска которых способствует поглощению излучения сине-фиолетовой части спектра, проникающего на эти глубины. Еще глубже, от 200 до 500 м, сила света ослабевает настолько, что фото-синтезирующие растения исчезают полностью.
Экологические области океана. В океане выделяют несколько областей с различными экологическими условиями: толщу воды - пелагиаль, среду обитания пелагических животных, и дно океана - бенталь, заселенную бентосом, т.е. организмами, обитающими на грунте или в толще грунта. Бенталь подразделяют на супралитораль, литораль, сублитораль, батиаль и абиссаль.
Супралитораль -побережье океана, расположенное выше уровня воды в самый высокий прилив, но еще подверженное эпизодическому воздействию океанических вод при нагонах и штормах. Здесь обитают как наземные, так и морские организмы.
Литораль -прибрежная зона морского дна, осушающаяся во время отлива. Располагается между отметками уровня воды в самый низкий отлив и в самый высокий прилив, простираясь до глубины 40 - 50 м. Обитатели литорали дважды в сутки покрываются водой и освобождаются от нее; они живут при условии резких изменений температуры и солености воды, действия прибоя, при прямом солнечном свете. Некоторые животные, прикрепляющиеся к скалам, могут поселяться так высоко, что покрываются водой лишь два раза в месяц, при максимальном сизигийном
приливе. Многие виды вне этой полосы существовать не могут, В приполярных областях, в полосе литорали жизнь крайне обеднена, так как значительную часть года она покрыта льдами.
Сублитораль -прибрежная часть океана, непосредственно примыкающая к литорали и располагающаяся на глубинах до 200 м, реже глубже, простирающаяся от уровня воды при самом низком отливе до нижней границы распространения донной растительности. С сублиторалью связаны богатые промыслы беспозвоночных.
Выделяются мелководья с мягким и твердым грунтом. На мягком грунте особенно обильны свойственные только мелководьям водоросли и цветковые растения. Из водорослей на более мелких участках преобладают зеленые и бурые, в том числе ламинарии и фукусы, а на больших глубинах - красные водоросли (багрянки). На мягком грунте мелководий обитают черви, крабы, офиуры, морские звезды, различные моллюски. Некоторые обитатели способны зарываться на глубину до 50 см, предпочитая песчаный грунт илистому, поскольку последний из-за обилия гниющих органических остатков характеризуется недостатком или отсутствием кислорода и заселен слабее песчаного.
На мелководьях со скалистым грунтом встречаются сверлящие организмы, активно внедряющиеся в твердый субстрат. Обитающие на поверхности субстрата усоногие раки и моллюски имеют крепкие раковины и прочно прикрепляются к грунту, что не позволяет волнам оторвать их. В пещерах и трещинах скальных грунтов поселяются головоногие моллюски, а также некоторые другие беспозвоночные и рыбы (например, мурены).
Сублитораль обычно прерывается предустьевыми пространствами с их сильно опресненной, а иногда даже пресной водой.
Хотя сублитораль, если ограничить ее площадь материковой отмелью до глубины 200 м, составляет всего 8 % площади океана (29 млн км2), она значительно богаче в фаунистическом и флористическом отношении по сравнению с остальными областями океана.
Батиаль располагается на материковых склонах на глубинах 200 - 2000 м между неритовой зоной и абиссалью. Эта область характеризуется слабой освещенностью в верхних частях и полным отсутствием видимых лучей спектра в нижних. Для нее характерны незначительные колебания температуры и солености. Здесь отлагаются преимущественно илы.
Абиссаль -область наибольших морских глубин, превышающих 2000 м, соответствующая ложу Мирового океана с чрезвычайно медленным передвижением воды от полюсов к экватору, постоянно низкими температурами (около 0°С), практически почти полным отсутствием света. Здесь не встречаются зеленые растения, накапливаются преимущественно органогенные (в том числе радиоляриевые и глобигериновые) илы (рис. 86).

Рис. 86. Распределение животного населения в море (Л. А. Зенкевич, 1951)
Толща воды - пелагиаль - подразделяется на эпипелагиалъ - освещенные слои воды до глубины 200 м и глубоководную пелагиаль. Для пелагиали характерны представители четырех групп организмов: нектона, планктона, плейстона и нейстона. Нектон - совокупность активно плавающих организмов, основные представители этой группы - пелагические рыбы. Виды планктона - организмы, переносимые морскими течениями и волнами, так как сами они могут передвигаться весьма медленно. Плейстон и нейстон - это небольшое число видов соответственно растений и животных, обитающих на поверхности воды.
Основу всей жизни пелагиали составляет фитопланктон. Малая скорость движения планктона ни в коей мере не может противостоять движению воды. У видов планктона (как и у представителей нектона) имеются приспособления, препятствующие погружению в толщу воды. Из них можно назвать следующие: уменьшение относительной плотности за счет выработки веществ, имеющих малую плотность (в первую очередь жиров), а также насыщения тканей и полостей тела водой (так, у некоторых медуз вода составляет 99 % массы тела). Отдельные виды имеют воздушные камеры, уменьшающие их плотность.
К фитопланктону относятся почти исключительно микроскопические одноклеточные организмы, имеющие хлорофилл и способные к ассимиляции. Они и являются источником первичной продукции. Значительно меньшую роль в этом отношении играют зеленые растения, прикрепленные ко дну, - различные водоросли и цветковые растения, а также хемотрофы.
Биомасса фитопланктона в Мировом океане составляет 1,5 млрд т, а его продукция - 559 млрд т. В этом состоит фундаментальное
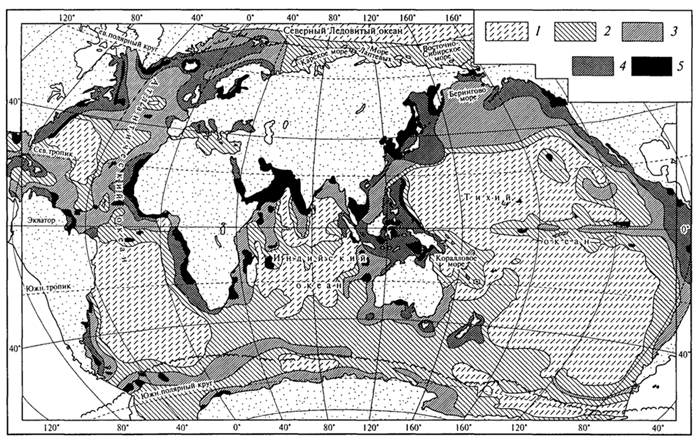
Рис. 87. Распределение средней годовой первичной продукции водорослей (мг/м2 в день) в Мировом океане (О.И.Кобленц-Мишке, 1985): 1- <100; 2- 100-150; 3-150-250; 4-250-500; 5 - свыше 500
отличие морских экосистем от наземных, где продукция составляет лишь небольшую часть общей биомассы. В целом водоросли в Мировом океане (включая фитопланктон) создают органическое вещество (сухая масса) в количестве примерно 74 % мировой продукции. "Урожайность" водорослей составляет 1,3 - 2,0 т сухого вещества с 1 га поверхности водной толщи в год (рис. 87).
Зоопланктон представлен микроскопическими и более крупными организмами: медузами и сифонофорами из кишечнополостных, сальпами и аппендикуляриями из хордовых.
Неосвещенная область океана, охватывающая батиаль и абиссаль, обладает некоторыми общими особенностями, связанными с пессимальными условиями существования.
Абиссаль занимает около 8 % морского дна и основную толщу морской воды, значительно превосходя по площади шельфовую область, а по объему - освещенную пелагиаль. При этом несомненно абиссальный бентос является производным от бентоса освещенной области океана, а население абиссальной пелагиали произошло от обитателей освещенной пелагиали.
Наряду с отсутствием света на отборе глубоководных форм сказываются высокое давление и низкая температура, характерные для глубин и создающие здесь пессимальные условия существования. Об уменьшении числа видов с глубиной свидетельствуют хотя бы данные океанологической экспедиции на корабле "Челленджер". Ею были обнаружены на разных глубинах следующее количество видов крабов: до 36 м - 190; 38 - 180 м - 75; 180 - 360 м - 28; 360 - 900 м - 21; 900- 1800 м - 3; 1800-3600 м - 2. Та же закономерность прослежена и для числа видов простейших - фораминифер, которое равно на глубинах до 100 м - 138, а на глубинах 5000 - 6000 м всего 19.








