Территория России и прилегающих стран относится, по мнению Л.С.Берга, к двум биогеографическим областям пресноводной фауны: Голарктической и Амурской переходной (бассейн р. Амура). В основу районирования Бергом был положен географический фактор - наличие изолированного водосборного бассейна с относящимися к нему реками и озерами. Такой подход полностью себя оправдал, что можно видеть, например, в новых схемах биогеографического районирования вод северо-востока Азии И.А.Черешнева (1992).
По Л.С.Бергу (1949), Голарктическая область включает всю Европу, Северную Африку, Азию к северу от Гималаев вместе с Передней Азией и Северную Америку с частью Центральной Америки. Только для этой области характерны некоторые семейства (лососевые, хариусы, корюшковые, щуковые, окуневые, умбровые, даллиевые) и роды рыб (налим, линь, красноперка и многие другие), миноги, а из ракообразных - речные раки, водяные ослики и др.
Голарктическая область была подразделена Л.С.Бергом на подобласти, из которых четыре приходятся на Евразию (рис. 96).
1. Циркумполярная (Кругополярная) подобласть включает бассейны всех рек, впадающих в Ледовитый океан и северную часть Тихого океана. Из рыб в ней резко преобладают лососевые. В общем фауна очень бедная, нет сомовых, усачей, сазанов и др., а из моллюсков отсутствуют Theodoxus, Melanopsis, Lithogliphus. Перловица живет только на западе подобласти.
2. Байкальская подобласть ограничена оз. Байкал, фауна которого описана выше. По мнению некоторых исследователей, это озеро настолько своеобразно, что должно быть поставлено вне
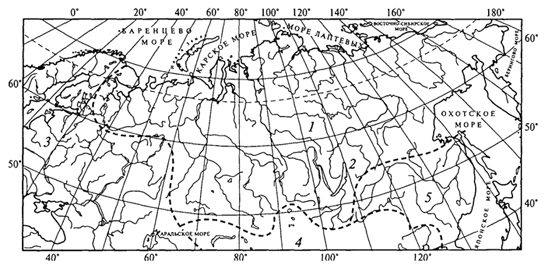
Рис. 96. Зоогеографическое деление пресноводных водоемов Северной Евразии (Л.С.Берг, 1933): Голарктическая область. Подобласти: 1 - Циркумполярная (Кругополярная); 2 -Байкальская; 3 -Средиземноморская; 4 -Нагорноазиатская; 5 - Амурская переходная
существующих зоогеографических категорий Голарктики и рассматриваться как самостоятельная биогеографическая область.
3. Средиземноморская подобласть занимает всю Европу, кроме северной ее части, входящей в Циркумполярную подобласть, Северную Африку, бассейны Черного, Каспийского и Аральского морей, северную часть Сирии и большую часть Ирана. Из рыб здесь преобладают карповые, лососевых сравнительно мало; характерны также сельдевые и бычки. Из моллюсков только в этой подобласти обитают многие роды, из ракообразных обыкновенные речные раки, пресноводные крабы и несколько родов каспийских бокоплавов. Многие характерные для подобласти формы происходят из солоноватых вод. Примечательна близость некоторых средиземноморских рыб к североамериканским и восточноазиатским. Лопатонос бассейна Арала близок к миссисипскому, умбра (евдошка) живет в Дунае, Днестре и восточной части США. Европейскую белугу в бассейне Амура замещает калуга.
4. Нагорноазиатская подобласть объединяет бассейны Центральной Азии, не имеющие стока в океан, бассейны Балхаша, а так же верховья Амударьи и Сырдарьи, Гильменда, Инда, Ганга, Брахмапутры, Меконга, Янцзы и Хуанхэ. Фауна бедная, но своеобразная. Из рыб характерны маринки, османы, гольцы, из моллюсков - особые виды прудовика, катушки и битинии.
5. Территорию России частично захватывает Амурская переходная область. К этой области относятся бассейны рек Амура, Тугура, Уды, а также о. Сахалин, Японские острова, Корейский полуостров,
бассейны рек, впадающих в Татарский пролив и Японское море. Она гораздо менее обширна, чем Голарктическая область, и называется переходной потому, что ее фауна наряду с эндемичными формами включает элементы двух соседних областей: лежащей севернее - Голарктики и находящейся южнее - Китайско-Индийской области. В Амуре обитают 85 видов рыб различного происхождения, в том числе много видов Средиземноморской подобласти, отсутствующих в Циркумполярной подобласти (белуга, вьюн, горчак, сазан, сом). Разрыв их ареалов связан с оледенением. Имеются также китайские формы (змееголов, желтощек, толстолобик). Следует отметить наличие в фауне бассейна Амура ряда беспозвоночных южного происхождения: креветок, некоторых губок, мшанок, моллюсков, водяных клещей.
В последние годы А. П. Андрияшевым, И. А. Черешневым, А.С.Новиковым, К.А.Савваитовой и другими детально изучена ихтиофауна северо-востока России. Я. И.Старобогатовым (1972) Палеарктика и Неарктика были повышены до ранга областей, что позволило И.А.Черешневу выделить переходную Берингийскую область на территории восточной части Чукотки, западной и арктической областей Аляски.
С применением биогеографического метода было обосновано предположение о слиянии в период регрессии моря в среднем плиоцене на Чукотском аваншельфе сибирских и аляскинских рек в единую речную систему, по которой происходило расселение пресноводных рыб между континентами. Завершение процесса формирования ихтиофауны приберенгийских территорий произошло после окончания последнего оледенения. При этом наиболее существенно изменился состав фауны в крупнейших реках Аляски - Маккензи и Юкон, куда вселилась большая группа рыб из южных районов Северной Америки через систему приледниковых озер. Таким же образом сибирские виды рыб заселили реки побережья Охотского моря.
Берингийская область удовлетворяет требованиям, традиционно предъявляемым к переходным зоогеографическим областям (Л.С.Берг, 1962), и характеризуется обитанием эндемичного таксона высокого ранга - семейства даллиевых рыб (три вида). Эндемичны также берингийский омуль, голец Андрияшева и реликтовая трехиглая колюшка. Своеобразен также состав остальной ихтиофауны (И. А. Черешнев, 1992).
ГЛАВА XII
БИОГЕОГРАФИЯ И ПРОБЛЕМЫ СОХРАНЕНИЯ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Среди проблем, с которыми сталкивается человечество, выделяется одна жизненно важная - это сохранение биологического разнообразия. Биота как связующее звено в системе Земля-Солнце является основным регулятором стабильности биосферы, источником пищи, сырьевых ресурсов, кислорода, чистой воды и воздуха. Мировое сообщество, встревоженное последствиями научно-технического прогресса, оказывающего зачастую разрушающее воздействие на условия существования биоты и в конечном итоге самого человека, уделяет много внимания проблемам биоразнообразия.
Понятие "биоразнообразие". Словосочетание "биологическое разнообразие" впервые применил энтомолог Г. Бэйтс (1892) в известной работе "Натуралист на Амазонке", когда описывал свои впечатления от встречи около 700 видов бабочек за время часовой экскурсии.
Понятие "биоразнообразие" в широком смысле охватывает множество различных биологических направлений и относится к разнообразию всего живого мира, являясь в какой-то мере синонимом понятия "жизнь на Земле". Оно используется активно не менее полувека. За это время много сделано для понимания самого явления и разработки методов его измерения. В научном мире биоразнообразие приложимо к таким фундаментальным понятиям, как гены, виды и экосистемы, которые соответствуют трем основным, иерархически зависимым уровням организации жизни на планете: генетическому, таксономическому и экологическому.
Попытки оценить биологическое разнообразие Земли предпринимались еще в XVIII-XIX вв., когда разрабатывались системы ботанико-географического и зоогеографического разделения поверхности планеты по своеобразию флоры и фауны. В XX в. сформировалось представление о разнообразии сообществ растений, животных и биомов планеты.
Одним из существенных достижений XX в. является расширение представлений о видовом разнообразии жизни на Земле. При всей неполноте наших знаний нельзя не отметить, что число таксонов животных и растений увеличено по меньшей мере в 500 раз по сравнению с концом XIX в. Если сейчас валидно описано 1,75 млн видов растений, животных, микроорганизмов, то, по мнению ведущих
специалистов-систематиков, реальное их число достигает не менее 10 - 35 млн, в том числе 1 млн видов микроорганизмов, 1 млн видов нематод, 10 млн видов насекомых и около 10 млн видов грибов - наименее изученных групп организмов. Среди биомов суши следует отметить влажные тропики, где, как полагают систематики, описан только один из 20 обитающих видов, особенно среди насекомых, грибов, а также представителей почвенной фауны.
УРОВНИ БИОРАЗНООБРАЗИЯ
В настоящее время биоразнообразие принято рассматривать как генетическое, популяционное видовое (таксономическое) и ценоти-ческое (сообщества и экосистемы).
Генетическое разнообразие и геногеография. Явление разнообразия жизни определяется фундаментальным свойством биологических макромолекул, особенно нуклеиновых кислот, - их способностью к спонтанным изменениям структуры, что приводит к изменениям геномов, к наследственной изменчивости. На этой биохимической основе разнообразие создается в результате трех независимо действующих процессов:
спонтанно возникающих генетических вариаций (мутаций),
естественного отбора в смешанных популяциях,
географической и репродуктивной изоляции.
Данные процессы, в свою очередь, ведут к дальнейшей таксономической и экологической дифференциации ценозов и экосистем.
До начала XX столетия объектами биогеографии были виды и более высокие таксоны животных и растений, их сообщества и экосистемы. Однако динамичные, быстро идущие процессы жизни самих видов в пределах их ареалов долгое время оставались слабо изученными. Только в 20-е годы XX в. внедрение в биогеографию идей и методов анализа популяций, популяционной генетики позволило начать изучение динамики вида, перейти к наблюдениям и картографированию динамичных биологических явлений, протекающих в масштабах времени, измеряемого продолжительностью жизни поколений - обычно это годы, для видов и экосистем - тысячи и миллионы лет. Генетические процессы в популяциях измеряются поколениями. От поколения к поколению происходят генетические изменения, которые и накапливаются в генофонде популяции. Вследствие этого генофонды различных популяций под действием медленных, но постоянных изменений генетически удаляются друг от друга.
Новой областью знаний, предметом которой является изучение географической изменчивости генофондов живой природы (в пределах ареала вида) на основе плодотворного синтеза методов географии, генетики и экологии, стала геногеография. Творцом
этого нового научного направления, как и автором самого термина "геногеография", появившегося в 1928 г., был замечательный русский генетик А.С.Серебровский (1892- 1948), известный не только своими работами, но и организацией первой кафедры генетики в Московском университете.
Доказано, что изменения, возникающие под действием естественного отбора, могут иметь разные последствия в зависимости от условий существования. В том случае, когда условия среды, определяющие естественный отбор, однородны на протяжении ареала, приспособленность вида к среде обитания неуклонно возрастает, а за достаточно продолжительный период времени таким путем могут возникнуть новые генотипы: генетическая структура отдельного ряда последовательных поколений постепенно и равномерно изменяется от поколения к поколению. Этот процесс называется филетической эволюцией.
Другой тип эволюционных преобразований наблюдается в том случае, когда разные популяции одного вида оказываются изолированными друг от друга в различных условиях среды. Поскольку при этом естественный отбор действует на них по-разному, изолированные популяции будут все более дивергировать, пока наконец единый исходный вид не распадется на два или более новых вида. Описанный процесс известен под названием видообразования. Этот процесс ведет к дроблению единой генетической популяции на ряд подгрупп, каждая из которых представляет собой независимую эволюционную линию. При этом дивергенция непрерывна: она не прекращается и после того, как данная группа достигла ранга вида, а продолжается и дальше, приводя к возникновению более высоких таксономических категорий.
Связь между видообразованием и географической изоляцией была замечена биогеографами еще в XIX в. Основываясь на идеях Ч.Дарвина, М. Вагнер в 1868 г. предложил свою "географическую" теорию видообразования, в которой содержалась идея о том, что географическая изоляция - необходимое условие образования видов из географически разобщенных рас и подвидов. Впоследствии эти представления были развиты Д.Джорданом, Б. Реншем, Ф. Г.Добжанским и Э.Майером. Дальнейшим развитием идей о географическом видообразовании стал географический популяционный анализ, основанный на математических моделях "островного" видообразования С.Райта (1943, 1946), на компьютерной обработке данных по генетической изменчивости популяций и видов в пределах ареала. Этот новый методический арсенал средств изучения популяций позволил упрочить позиции геногеографии или генетической биогеографии как активно развивающейся области биогеографической науки.
Генетики выделили удобный объект для сравнения организмов, каковым является геном - совокупность генов, характерных
для гаплоидного набора хромосом любого организма. Геном - функциональная единица, своего рода программа, необходимая для нормального развития и воспроизводства организма в ряду поколений. Современная генетика располагает многочисленными средствами исследования геномов - геномным анализом.
В разнообразии геномов домашних животных А. С. Серебровский еще в 20-е годы XX в. призывал видеть естественное богатство и обозначил этот генетический запас термином "генофонд". Основной задачей геногеографии стало изучение пространственного распространения генофонда. А. С. Серебровский (1928) писал: "Изучение географического распространения концентраций различных генов и должно составить то, что мы предлагаем назвать геногеографией".
Советские генетики в 20 - 30-х годах не только заложили основы геногеографии, но и дали блестящие образцы таких исследований, выполненных в разном географическом масштабе. Среди них особое место занимают ставшие классическими работы Н.И.Вавилова по мировой геногеографии культурных растений.
География распространения генов в ареале вида является пространственным воплощением генетических процессов, а географическое картографирование генофонда - способом увидеть его в зримых картографических образах, разглядеть процессы, протекающие в генофонде, уловить в нем и универсальные для всех популяций черты, и те особенные черты, что отличают генофонд каждой из них от всех других На основе этих данных можно построить карту географического распределения генов, или геногеографическую карту - основной инструмент в работе геногеографа (рис. 97).
Генетика и экология популяций. В генетическом смысле популяция - это пространственно-временная группа особей одного вида, в которой в течение достаточно длительного времени осуществляется свободное скрещивание - панмиксия. Для осуществления панмиксии необходимо отсутствие изоляционных барьеров для скрещивания. Разные же популяции одного вида изолированы друг от друга именно такими барьерами, или препятствиями для свободного скрещивания, свободного обмена генетической информацией.
Известный датский генетик В.Иогансен в 1903 г. предложил различать "популяции" - генетически неоднородные группы особей одного вида и "чистые линии" -генетически однородное потомство одной особи.
Вследствие репродуктивной связи между поколениями генетическая популяция непрерывна во времени, а внутрипопуляционные скрещивания обеспечивают ей пространственное единство. Численность популяции может возрастать или уменьшаться в результате миграции особей, а также изменения показателей рождаемости и смертности. Популяция может объединиться с другими

Рис. 97. Распространение зебр и клинальная изменчивость полосатости (Л.Миттлер, Т.Грегг, 1972): 1 - зебра Греви; 2 -зебра Гранта; 3 -зебра Чэтмена; 4 -бурчеллева зебра; 5 - квагга; 6 -зебра Гартмана; 7 -капская горная зебра. Распространение зебры в областях, расположенных ниже пунктирной линии, основано на старых данных о формах, теперь уже вымерших; зебра Гранта, зебра Чэтмена и бурчеллева зебра обнаруживают клинальную изменчивость полосатости; эти формы представляют собой расы одного многомерного вида, отличного от зебры Греви и горной зебры
популяциями или исчезнуть вследствие эмиграции всех ее членов, а также вымирания.
Степень панмиксии внутри популяции у разных видов может значительно колебаться по разным причинам. Так, у одних видов особи образуют пары на всю жизнь (например, лебеди), у других - только на сезон размножения (например, многие утки), для третьих - характерно образование нестойких пар (тетерева и глухари), у четвертых - самки оплодотворяются только один раз на протяжении всей жизни (многие насекомые, паукообразные),
наконец, есть виды с наружным оплодотворением (большинство гидробионтов), у которых при наличии массовых нерестилищ оплодотворение группы яйцеклеток может осуществляться смесью сперматозоидов от разных особей. Все эти способы оплодотворения, конечно, влияют на степень панмиксии, присущую всем популяциям видов.
Выделение популяций нередко оказывается затруднительным. Границы между ними часто бывают стертыми, наблюдается постепенное изменение признаков в том или ином географическом направлении. Такой характер географической изменчивости генофонда получил название клинальной изменчивости, впервые описанной Дж. Хаксли в 1939 г. (рис. 98). В настоящее время разработаны методы использования разнообразных, особенно генетических и биохимических, маркеров популяций, что без труда позволяет дифференцировать популяции внутри видовых ареалов.

Рис. 98. Относительные частоты неполосатых (белые части кружков) и полосатых (черные части кружков) улиток в ряде соприкасающихся колоний, обитающих вдоль реки Арьеж во французских Пиренеях (Л. Миттлер, Т. Грегг, 1972)
На образование популяций растений и животных сильнейшее воздействие оказывают особенности географической среды, ее ландшафтное и фациальное разнообразие, иначе говоря, разнообразие биотопов, которые может заселить вид. Существуют определенные различия между популяциями животных и растений, поскольку растения в большей степени зависят от локальных особенностей среды обитания и отличаются большей изменчивостью. В образовании популяций у растений определяющее значение имеют особенности размножения, позволяющие поддерживать генетическое единство популяций: дальность распространения семян и пыльцы, семенную продуктивность, способность вида к изменчивости и скрещиванию с близкими видами.
В качестве изолирующих факторов прежде всего рассматриваются орографические особенности территории, наличие крупных поднятий, препятствующих переносу семян и пыльцы растений и расселению животных. Некоторое изолирующее значение
могут иметь и крупные реки. Серьезным препятствием для контакта между популяциями являются пространства, где данный вид отсутствует. Так, для популяций лесных растений изолирующее значение имеют крупные болотные массивы или обширные степные пространства, разделяющие, например, популяции сосны обыкновенной в Северном Казахстане.
Роль барьеров могут играть и факторы биотической среды. Так, уничтожение, естественная гибель или миграции птиц, распространяющих семена некоторых растений, могут способствовать распаду единой популяции. Значение биотических изоляторов усиливается в связи с антропогенным воздействием на природу. Установлено, что у многих видов внутри популяции население распадается на небольшие и сравнительно изолированные группки особей - демы (от греч. "демос" - народ), степень панмиксии в которых, естественно, оказывается выше, чем во всей популяции. Известно, что демы, во-первых, относительно кратковременные и нестойкие объединения особей, существующие, например, у мелких мышевидных грызунов одно-два поколения. Такие мелкие внутрипопуляционные группировки всегда бывают немногочисленными и не могут рассматриваться как самостоятельные образования (Н.В.Тимофеев-Ресовский, А.В.Яблоков, 1974).
Важная характеристика каждой популяции - численность составляющих ее особей, которая также может быть очень различной. С одной стороны, популяции некоторых насекомых включают многие миллионы особей, с другой - популяции многих крупных животных могут включать лишь сотни особей. Внутри каждого вида бывают и мелкие, и крупные популяции. Так, прыткие ящерицы в одних случаях образуют популяции, состоящие из нескольких сотен особей, в других - из многих тысяч особей (рис. 99). К тому же многим видам свойственны резкие колебания численности популяций. Например, в популяции кроликов на островке Скокольм около Англии, куда они были завезены почти 600 лет назад и хорошо прижились, на протяжении нескольких лет численность изменялась от 10 000 до 100 особей.
Популяция - группа динамичная, как и входящие в нее особи, она рождается, живет, умирает. При этом рождаемость, продолжительность жизни и смертность особей составляют важнейшие характеристики любой популяции, которая, как было замечено еще Ч.Дарвином, всеми силами стремится к повышению своей численности и максимальному распространению. Подсчитано, что одна диатомовая водоросль могла бы в течение восьми дней дать массу вещества, по объему равную Земле, а в течение следующего одного часа удвоить эту массу. Даже самые медленно размножающиеся животные - слоны - могли бы очень быстро завоевать всю Землю, будь для этого благоприятные условия: по подсчету Дарвина, от одной пары слонов за 750 лет получилось бы 19 млн голов
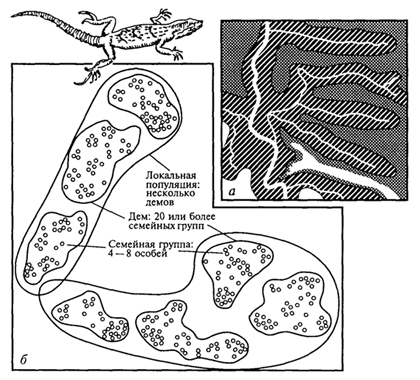
Рис. 99. Распространение прыткой ящерицы внутри ареала (А. В.Яблоков, 1987): а -сгущения плотности населения ящериц вдоль рек и ручьев; б -демы и семейные группы внутри демов
потомства, а за 3000 лет слоны покрыли бы сплошным слоем всю поверхность земного шара. Ничего подобного в реальности не происходит, так как для беспрепятственного размножения у видов условий нет, но всегда есть ограниченность пищевых ресурсов, пригодных для жизни мест, воздействие болезней, врагов и конкурентов.
Таким образом, несмотря на значительные различия в площади, численности, структуре и т.п., популяции являются реально существующими группировками особей, на которые распадаются любые виды живых организмов. Это позволяет считать популяцию реальной элементарной единицей эволюционного процесса.
Микро- и макроэволюция. Микроэволюция - эволюция в популяциях под влиянием ненаправленной мутационной изменчивости на внутривидовом уровне, когда особи способны скрещиваться и давать плодовитое потомство. Термин "микроэволюция" ввел в научный обиход генетик Ю. А. Филипченко в 1927 г., чтобы разграничить два принципиально важных эволюционных явления. Макроэволюция -процесс развития на уровнях выше видового обмена генетической информацией между особями разных видов, но четко проявляются тенденции адаптации животных и растений к биотическим и абиотическим факторам окружающей среды.
В.Йогансен в 1909 г. ввел разграничение между фенотипом и генотипом. Фенотип -это совокупность всех внешних наблюдаемых признаков организма, будь то морфологические, физиологические или поведенческие признаки. Генотипом называется совокупность всех наследуемых задатков данной клетки или организма ("генетическая конституция" особи). На протяжении жизни организма его фенотип может изменяться, однако генотип остается неизменным.
Эволюцию на генетическом уровне можно рассматривать как двухступенчатый процесс. С одной стороны, возникают мутации и рекомбинации - процессы, обусловливающие генетическую изменчивость; с другой - наблюдается дрейф генов и естественный отбор - процессы, посредством которых генетическая изменчивость передается из поколения в поколение. Эволюция возможна только в том случае, если существует наследственная изменчивость. Единственным источником новых генетических вариантов служит мутационный процесс, но генетические варианты, возникшие в результате мутационного и рекомбинационного процессов, передаются из поколения в поколение отнюдь не с равным успехом: частота некоторых из них может увеличиваться за счет других. Помимо мутаций к процессам, изменяющим частоты аллелей в популяции, относятся естественный отбор, поток генов (т.е. миграция их) между популяциями и случайный дрейф генов.
При изучении процесса эволюции важное значение имеет представление о генофонде - совокупности генотипов всех особей популяции. Для диплоидных организмов генофонд популяции, насчитывающей N особей, состоит из 2N гаплоидных геномов. Каждый геном содержит всю генетическую информацию, полученную организмом от одного из родителей. Процессы, изменяющие частоты генов, постоянно происходят в популяциях, и без них не было бы эволюции.
С.С.Четвериков обратил внимание на то, что, поскольку любая природная популяция имеет ограниченную численность особей, это неизбежно приведет к чисто случайным, статистическим процессам в распространении мутаций. Численность популяций всех видов постоянно колеблется (например, у грызунов в лесу она может изменяться в сотни, а у многих видов насекомых - в десятки тысяч раз), из-за чего в разные годы распространение мутаций в популяциях может идти по-разному. От громадной популяции птиц, насекомых, зайцев или других животных в трудный для переживания год может остаться всего несколько особей, причем иногда не типичных для бывшей популяции. Но именно они передадут потомству свой генофонд, поэтому новая популяция по составу генетического материала будет иной, чем прежняя. В этом проявляется генетический "эффект основателя популяции".
Непрерывно изменяется и геном в популяциях человека. К. Аль-стрем на материале по Южной Швеции показал, что в популяции человека передается следующему поколению далеко не весь имеющийся генофонд, а лишь избранная, а то и случайно "выхваченная" часть. По его данным, 20 % поколения здесь вовсе не оставили потомков, зато 25 % родителей, которые имели трех и более детей, дали 55 % численности следующего поколения.
Американский генетик и селекционер И. М.Лернер в 1954 г. сформулировал понятие о генетическом гомеостазе, определив его как способность популяции приводить в равновесие свою генетическую структуру и противостоять внезапным изменениям. Один из важных механизмов генетического гомеостаза - отбор в пользу гетерозигот, ведущий к сбалансированному равновесию.
Постоянное давление мутаций и миграции генов, а также выщепление биологически менее приспособленных генотипов создают проблему так называемого генетического груза. Это понятие ввел лауреат Нобелевской премии Г. Меллер в 1950 г. По его расчетам, от 10 до 50 % гамет у человека содержат хотя бы одну вновь возникшую мутацию. Слабо вредящие мутации, если только они проявляются в гетерозиготе, способны нанести популяции больший урон, чем полностью рецессивные летальные мутации. Г. Меллером с соавторами (1956) предложены расчеты летального эквивалента, соответствующего числу мутаций, дающих вместе летальный исход. Было показано, что средняя величина "генетического груза" у человека равна 3 - 5 летальным эквивалентам.
Новые аллели могут поступать в генофонд популяции в результате мутаций, а также в виде гамет, привнесенных иммигрантами из других популяций. Процесс поступления "чужих" генов называется обменом генами, или, если это происходит постоянно, потоком генов. Мутационный процесс и обмен генами сходны в том отношении, что оба они поставляют популяции новые или дополнительные аллели, т.е. оба служат источником изменчивости.
Демы, географические изоляты, расы и даже виды очень редко представляют собой замкнутые системы. Обычно между ними всегда происходит некоторый обмен генами, причем чем теснее пространственная и генетическая близость популяций, тем больше поток генов.
Представления о существенных различиях между генетически эффективной и общей численностью популяции были развиты теоретически и при экспериментальном изучении стохастических процессов изменения генных частот. Такого рода "случайный дрейф генов" - математический факт, вытекающий из явления конечности численности любой реальной популяции.
Полиморфизм -проявление индивидуальной прерывистой изменчивости живых организмов: наличие в одной и той же популяции
двух или более хорошо обозначенных форм, способных появляться в потомстве одной самки и встречающихся с частотой, достаточно высокой для того, чтобы исключить поддержание самой редкой из них повторяющимися мутациями. Обычно на практике полиморфной считают популяцию с частотой гетерозигот по некоторому локусу более 1 - 5%. Анализ полиморфных признаков - ключ к изучению генетических процессов в популяциях, в связи с чем ранние исследования такого рода были выполнены на видах с хорошо выраженным полиморфизмом внешнего облика особей. Накоплено множество данных о феноменологии, генетическом контроле и механизмах поддержания полиморфизма у разных видов.
Наряду с очевидным полиморфизмом практически у любого вида существует огромный запас скрытой наследственной изменчивости. Генетическая гетерогенность -это любое, пусть даже самое минимальное генетическое разнообразие в популяции, вызываемое процессом мутации или рекомбинации. Полиморфизм, как следует из только что данного определения, есть устойчивое поддержание в популяции достаточно многочисленных генотипических классов, что возможно лишь при участии отбора.
Концентрация мутаций есть показатель, характеризующий популяцию в целом. Он устанавливается под действием генетико-автоматических процессов и отбора. Разные популяции отличаются друг от друга по количеству и составу мутаций. Эти показатели со временем претерпевают изменения в результате относительных вкладов разных генотипов из размножающейся части популяции в генофонд следующего поколения за счет их дифференциального воспроизведения или выживаемости.
Таким образом, вид представляет собой сложное образование. Он распадается на целую систему рас и популяций, которые, в свою очередь, слагаются из биологически неравноценных особей.
Рассмотрим некоторые примеры. Как известно, в популяциях большинства видов моллюсков встречаются особи с прямо противоположными по закрученности раковинками. Если у типичных особей данного вида раковинка закручена направо, по ходу часовой стрелки, то у уклоняющихся форм она закручена налево, против часовой стрелки. Частота уклоняющихся раковинок может быть очень различной у разных видов и в разных популяциях одного и того же вида. В. Н. Цветков (1938) изучил распространение право- и левозавитковых форм у моллюсков, обитающих в окрестностях Алма-Аты (рис. 100). Здесь встречаются преимущественно правозавитковые особи. Ограничение распространения левозавитковых форм, по-видимому, обусловлено тем, что они по сравнению с правозавитковыми обладают большей интенсивностью обмена веществ и меньшей способностью к ассимиляции пищи. Эти особенности лимитируют распространение левозавитковых форм
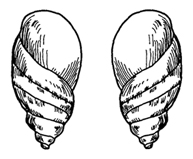
Рис. 100. Правозавитковая (дектральная) и левозавитковая (синистральная) раковинки у моллюсков (Н.П.Дубинин, 1966)
в Семиречье, так как здесь из-за частых и длительных засух регулярно наступают периоды голодания моллюсков.
В процессах экологической дивергенции популяций на первый план выступает адаптация к локальным условиям среды. Как указывал В. Н. Сукачев (1928), популяции сосны представлены рядом экологических рас соответственно условиям обитания. Чем южнее происхождение сосны, тем хуже она переносит низкую температуру. Так, температура, безвредная для скандинавского экотипа, губительна для среднеевропейских сосен. Оказалось, что основная причина этих различий - биохимические особенности тканей деревьев.
Популяции - первая ступень, где разыгрываются явления эволюции. Зачинаясь в отдельных популяциях, процессы эволюции лишь в конечной форме, пройдя через многообразные ступени, обусловленные сложной динамической внутривидовой системой, завершаются актом происхождения нового вида.
Только небольшая часть (около 1 %) генетического материала высших организмов изучена в достаточной мере, когда мы знаем, какие гены отвечают за те или иные проявления фенотипа организмов. Для большей части ДНК ее значение для вариации жизненных форм остается неизвестным.
Каждый из 109 различных генов, распределенных в мировой биоте, не дает идентичного вклада в формирование разнообразия. В частности, гены, контролирующие фундаментальные биохимические процессы, являются строго консервативными у различных таксонов и в основном демонстрируют слабую вариабельность, которая сильно связана с жизнеспособностью организмов.
Если судить об утере генофонда, принимая во внимание то, что каждая форма жизни уникальна, вымирание всего лишь одного дикого вида означает безвозвратную потерю от тысячи до сотен тысяч генов с неизвестными потенциальными свойствами. Генная инженерия могла бы использовать это разнообразие для развития медицины и создания новых пищевых ресурсов. Однако разрушение местообитаний и ограничение размножения многих видов приводят к опасному уменьшению генетической изменчивости, сокращая их способности адаптироваться к загрязнению, изменениям климата, болезням и другим неблагоприятным факторам. Основной резервуар генетических ресурсов - природные экосистемы - оказался значительно измененным или разрушенным. Уменьшение генотипического разнообразия, происходящее
под воздействием человека, ставит на грань риска возможность будущих адаптации в экосистемах.
Биологическая эволюция - это процесс накопления изменений в организмах и увеличение их разнообразия во времени. В основе всех этих изменений лежат генетические изменения, т.е. изменения наследственного вещества, которое, взаимодействуя со средой, определяет все признаки организмов. На генетическом уровне эволюция представляет собой накопление изменений в генетической структуре популяций.
Видовое разнообразие организмов. Разнообразие видов обычно оценивается исходя из внутривидовой (вид - подвид - популяции разного уровня) и надвидовой (род - семейство - до царства) таксономии. Видовой уровень разнообразия обычно рассматривается как базовый, а вид является опорной единицей учета биоразнообразия.
Р.Уиттекер, К.Виллис, Р.Филд (R.Whittaker, K.Willis, R.Field, 2001), развивая концепцию биологического разнообразия, подчеркивают, что она основана на признании не только феномена богатства видов, но также и биогеографических причин, его обусловливающих, в том числе большое значение придается историческому фактору и пространству, в котором формируется разнообразие. В процессе разработки этой теории сложилось представление о базовых единицах биоразнообразия.
Альфа-разнообразие -разнообразие видов (видовое богатство), которое обычно выражается числом видов животных или растений на единицу площади в определенной стандартной выборке, часто внутри одного сообщества или местообитания (локальный уровень).
Бета-разнообразие -разнообразие видов в сообществах по градиентам факторов среды (по катене) в пределах ландшафта; это определенная степень различия в видовом составе между разными типами сообществ или местообитаний (ландшафтный уровень).
Гамма-разнообразие -разнообразие видов в пределах крупных регионов в соответствии с дифференциацией условий по важнейшим г








