Процесс отщепления карбоксильной группы аминокислот в виде СО2 получил название декарбоксилирования. Несмотря на ограниченный круг аминокислот и их производных, подвергающихся декарбоксилиро-ванию в животных тканях, образующиеся продукты реакции – биогенные амины – оказывают сильное фармакологическое действие на множество физиологических функций человека и животных.
В живых организмах открыты 4 типа декарбоксилирования аминокислот:
1. α-Декарбоксилирование, характерное для тканей животных, при котором от аминокислот отщепляется карбоксильная группа, стоящая по соседству с α-углеродным атомом. Продуктами реакции являются СО2 и биогенные амины:

2. ω-Декарбоксилирование, свойственное микроорганизмам. Например, из аспарагиновой кислоты этим путем образуется α-аланин:

3. Декарбоксилирование, связанное с реакцией трансаминирования:
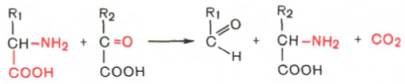
В этой реакции образуются альдегид и новая аминокислота, соответствующая исходной кетокислоте.
4. Декарбоксилирование, связанное с реакцией конденсации двух молекул:

Эта реакция в тканях животных осуществляется при синтезе δ-амино-левулиновой кислоты из глицина и сукцинил-КоА (см. главу 13) и при синтезе сфинголипидов, а также у растений при синтезе биотина.
Реакции декарбоксилирования в отличие от других процессов промежуточного обмена аминокислот являются необратимыми. Они катализируются специфическими ферментами – декарбоксилазами аминокислот, отличающимися от декарбоксилаз α-кетокислот (см. главу 10) как белковым компонентом, так и природой кофермента. Декарбоксилазы аминокислот состоят из белковой части,обеспечивающей специфичность действия, и простетической группы, представленной пиридоксальфосфатом (ПФ), как и у трансаминаз.
Таким образом, в двух совершенно различных процессах обмена аминокислот участвует один и тот же кофермент. Исключение составляют две декарбоксилазы: гистидиндекарбоксилаза Micrococcus и Lactobacilus и аденозилметионин-декарбоксилаза Е. coli, содержащие вместо ПФ остаток пировиноградной кислоты.
Механизм реакции декарбоксилирования аминокислот в соответствии с общей теорией пиридоксалевого катализа (см. рис. 12.3) сводится к образованию ПФ-субстратного комплекса, представленного, как и в реакциях трансаминирования, шиффовым основаниемПФ и аминокислоты:

Образование подобного комплекса в сочетании с некоторым оттягиванием электронов белковой частью молекулы ферментасопровождается лабилизацией одной из трех связей при α-углеродном атоме, благодаря чему аминокислота способна вступать вреакции трансаминирования (а), декарбоксилирования (b) и альдольного расщепления (с).
Билет 55
Эритроциты примерно через 120 дней разрушаются (лизируются) в кровеносном русле или в селезенке. Гемоглобин, освобождающийся из эритроцитов в крови, связывается гаптоглобином (специальный а2-глобулин плазмы) и в виде комплекса гемоглобин — гаптоглобнн попадает в клетки ретикулоэндотелиальной системы (РЭС), главным образом селезенки. Гемоглобин окисляется в метгемоглобин (Fe**"), а затем подвергается распаду. При этом гаптоглобнн отщепляется.и переходит в кровь. В клетках РЭС происходит первая фаза распада гемоглобина, сопровождающаяся образованием из него билирубина: 

Сначала под действием гемоксигеназы происходит окислительное расщепление а-метинового мостика гема, соединяющего два соседних пиррольных кольца. Кольцевая структура гема разрывается и образуется вердоело-бин. Последний теряет железо, которое связывается белком-переносчиком трансферином и доставляется с кровью в костный мозг и глобин. Глобин гидролнзуется катепсинами селезенки до аминокислот. Разомкнутая тетра- лиррольная цепь вердоглобнна называется билидердином (желчный пигмент зеленого цвета). При восстановлении биливердиыа образуется билирубин — пигмент красно-желтого цвета.
Билирубин — плохо растворимое в воде соединение, поэтому, поступая в Кровь, он связывается белком плазмы альбумином. Комплекс альбумин — билирубин — важнейшая нормальная транспортная форма желчных пигментов. С током крови билирубин поступает в клетки печени. Как липофильное вещество он легко проникает'через мембрану клеток печени, освобождаясь от альбумина.
Внутри клеток печени происходит вторая фаза превращения гемпротеидов (рис. 61). В печейн образуются конъюгированные формы билирубина — билирубинглюкурониды. Донором глюкуроновой кислоты выступает УДФ-глю-куроновая кислота. Реакция катализируется УДФ^люкуронозилтрпнсферй-зой. Образуются билирубннмоноглюкуронид (20%) и билирубинднглюкуро-нид (80%). Билирубинглюкурониды — хорошо растворимые в воде соединения.
Билирубинглюкурониды лишь в незначительных количествах могут диффундировать в кровеносный капилляр. Поэтому в плазме крови присутствуют две формы билирубина: некокъюгированный (он же непрямой, или свободный) и конъюгированный (он же прямой, или связанный). На долю первого приходится около 75% общего билирубина плазмы крови, на долю второго — около 25%.
Билирубинглюкурониды выделяются с желчью в кишечник, где происходит заключительная, третья фаза распада гемпротеидов. В желчных путях отщепляется гликуроновая кислота от билирубинглюкуроннлов и вновь образуется неконъюгнрованныЙ билирубин. В тонком кишечнике небольшая часть билирубина может всасываться и через портальную вену вновь поступать в печень, а оттуда опять с желчью выделяться нз кишечника. Эта. печеночно-кишечная циркуляция, сходная с циркуляцией желчных кислот, возможно, имеет биологическое значение. Большая же масса билирубина подвергается многократному восстановлению бактериями кишечника или редуктазамн слизистой кишечника. В тонком кишечнике билирубин превращается в мезобилирубин, затем мезобилирубиноген (или уробилиноген). Последний всасывается в тонком кишечнике и через воротную вену поступает в печень, где уробилиноген необратимо разрушается до моно- и дипирролов.Неразрушенный уробилиноген вновь поступает с желчью в кишечник. В толстом кишечнике мезобилирубиноген (уробилиноген) восстанавливается анаэробными бактериями до стеркобилииогена, который, как и уробилиноген, бесцветен. Большая часть стеркобилииогена выделяется с фекалиями и быстро окисляется кислородом воздуха до стеркобилина — оранжево-желтого пигмента, в основном определяющего цвет фекалий.
Небольшие количества стеркобилииогена всасываются в прямой кишке и через- систему геморроидальных вен, минуя печень, достигают с током крови почек, которые выводят его с мочой, Стеркобилиноген мочи, как и в фекалиях, окисляется в стеркобилнн, частично определяя нормальный соломенно-желтый Цвет мОчи, Ранее считалось, _что с мочой выделяетсЯ-уробнлн-ноген (это нашло отражение в его названии), который окисляется в уробилин, имеющий ту же окраску, что и стеркобилин. В норме уробилиноген не выделяется ни с мочой, ни с фекалиями.
Итак, в норме промежуточный продукт распада гема — билирубин — не накапливается в крови, а быстро захватывается клетками печени. По содержанию билирубина и соотношению его форм в крови можно судить, с одной стороны, о скорости распада гемпротеидов в клетках РЭС, а с другой — о превращении билирубина в печени. В норме содержание общего билирубина в сыворотке крови составляет 8—20 мкмоль/л (из них 75% неконъюгированного).
Конечный продукт превращения билирубина — стеркобилиноген — выделяетсяу человека в основном с калом (примерно 300 мг в сутки) и в незначительных количествах с мочой (около 1—4 мг в сутки). В норме билирубин в моче практически отсутствует (нижний предел чувствительности используемых методов позволяет определить конъюгированный билирубин в моче при его концентрации свыше 0,3 мг/л). Непрямой (неконъюгированный) билирубин может попасть в мочу только, если его повышение в сыворотке крови сочетается с нарушением проницаемости гломерулярной мембраны. Прямой конъюгированный билирубин в плазме крови в норме отсутствует. При патологии элиминация прямого (конъюгированного) билирубина с мочой значительно варьирует. При остром вирусном гепатите увеличение концентрации прямого билирубина в моче может быть в дожелтушный период, что существенно при массовом обследовании (диспансеризации) на гепатит. При выздоровлении билирубин исчезает из мочи. Патологическая билирубинурия может иметь место при следующих заболеваниях: инфекционный гепатит, цирроз печени, заболевания желчного пузыря и желчных протоков, метастатическая карцинома печени или гематома, токсическое повреждение печени. Под влиянием различных факторов в организме может нарушаться образование, превращение и выведение билирубина. Повышение содержания билирубина в крови ведет к отложению его в тканях, в том числе в коже и слизистых, и вызывает окрашивание их в коричневато-желтый цвет (цвет билиру бина). Эти состояния называют желтухами, которых бывает несколько видов: гемолитическая (или надпеченочная), паренхиматозная (или гепатоцеллю-лярная) и обтурационная (или подпеченочная). 
Билет 56
Переваривание нуклеопротеинов и всасывание продуктов их распада осуществляются в пищеварительном тракте. Под влиянием ферментов желудка, частично соляной кислоты, нуклеопротеины пищи распадаются на полипептиды и нуклеиновые кислоты; первые в кишечнике подвергаются гидролитическому расщеплению до свободных аминокислот. Распад нуклеиновых кислот происходит в тонкой кишке в основном гидролитическим путем под действием ДНК- и РНКазы панкреатического сока. Продуктами реакции при действии РНКазы являются пуриновые и пи-римидиновые мононуклеотиды, смесь ди- и тринуклеотидов и резистентные к действию РНКазы олигонуклеотиды. В результате действия ДНКазы образуются в основном динуклеотиды, олигонуклеотиды и небольшое количество мононуклеотидов. Полный гидролиз нуклеиновых кислот до стадии мононуклеотидов осуществляется, очевидно, другими, менее изученными ферментами (фосфодиэстеразами) слизистой оболочки кишечника.
В отношении дальнейшей судьбы мононуклеотидов существует два предположения. Считают, что мононуклеотиды в кишечнике под действием неспецифических фосфатаз (кислой и щелочной), которые гидролизируют фосфоэфирную связь мононуклеотида («нуклеотидазное» действие), расщепляются с образованием нуклеозидов и фосфорной кислоты и в таком виде всасываются. Согласно второму предположению, мононуклеотиды всасываются, а распад их происходит в клетках слизистой оболочки кишечника. Имеются также доказательства существования в стенке кишечника нуклеотидаз, катализирующих гидролитический распад моно-нуклеотидов. Дальнейший распад образовавшихся нуклеозидов осуществляется внутри клеток слизистой оболочки преимущественно фосфороли-тическим, а не гидролитическим путем.
Всасываются преимущественно нуклеозиды, и в таком виде часть азотистых оснований может быть использована для синтеза нуклеиновых кислот организма. Если происходит дальнейший распад нуклеозидов до свободных пуриновых и пиримидиновых оснований, то гуанин не используется для синтетических целей. Другие основания, как показывают опыты с меченными по азоту аденином и урацилом, в тканях могут включаться в состав нуклеиновых кислот. Однако экспериментальные данные свидетельствуют, что биосинтез азотистых оснований, входящих в состав нуклеиновых кислот органов и тканей, протекает преимущественно, если не целиком, de novo из низкомолекулярных азотистых и безазотистых предшественников.
Нуклеиновые кислоты гидролнзуются в тканях организма с помощью нуклеаз — деэокснрибоиуклеаз (ДНКазы) и рибонуклеаз (РНКаэы). Эти ферменты катализируют расщепление внутренних илн концевых межнуклеотидных связей (3', У-фосфодиэфирных) в ДНК и РНК. Иуклеазы, расщепляющие связи внутри полннуклеотидной цепи, называются эндонуклеазами, а отщепляющие концевые нуклеотиды — экзонуклеазами. Некоторые экзокуклеезы способны парализовать как ДНК, так и РНК. Под действием нуклеаз поли-нухлеотидная цепь распадается до олигонуклеотидов и мононуклеотидов (нуклеознд-5'- или З'-монофосфатов).

Мононуклеотиды распадаются до конечных продуктов обмена -гидролити-ческнм или фосфоролнтическим путями (в первом случае для разрыва связей используется вода, а во втором — фосфорная кислота),
Моновукдеотиды расщепляются с помощью нуклвотидаз {5'-нуклеотн-дазы и. З'-нуклеотндазы) и нуклеозидгидролвз до свободных оснований, пентозы и фосфата.
Пуриновые основания окисляются до мочевой кислоты: 
Она является конечным продуктом обмена пуриновых нуклеотидов в организме человека и выделяется с мочой. Превращение пирнмидиновых оснований идет до р-аланина, СОя и аммиака. Р-Аланин используется для синтеза дипептидов мышц — карноз'ина и ансерина — или выделяется с мочой.
Таким образом, содержание мочевой кислоты в крови и моче отражает интенсивность расщепления нуклеиновых кислот в организме.
Нарушения обмена пуринов
Подагра
Когда гиперурикемия принимает хронический характер, говорят о развитии подагры (греч. poclos – нога, agra – захват, дословно – "нога в капкане").
В крови мочевая кислота находится в форме ее солей – уратов натрия. Растворимость уратов в плазме крови невелика и при превышении порога растворимости в плазме (около 0,7 ммоль/л) они кристаллизуются в периферических зонах с пониженной температурой, образуя тофусы (греч. tophus – пористый камень, туф). Накапливающиеся в межклеточном веществе ураты некоторое время фагоцитируются, но фагоциты не способны разрушить пуриновое кольцо. В результате это приводит к гибели самих фагоцитов, к выходу лизосомальных ферментов, активации свободнорадикального окисления и развитию острой воспалительной реакции – развивается подагрический артрит. В 50-75% случаев первым признаком заболевания является мучительная ночная боль в больших пальцах ног.
Мочекаменная болезнь
Мочекаменная болезнь заключается в образовании солевых кристаллов (камней) разной природы в мочевыводящих путях. Непосредственно образование мочекислых камней составляет около 15% от всех случаев этой болезни. Мочекислые камни в мочевыводящих путях откладываются примерно у половины больных подагрой.
|
Две переходные формы мочевой кислоты при pH 5,75
|
Наиболее часто такие камни представлены в дистальных канальцах и собирательных трубочках. Причиной отложения кристаллов мочевой кислоты является гиперурикемия и повышенное выведение уратов натрия с мочой. Главным провоцирующим фактором кристаллизации является увеличение кислотности мочи. При понижении рН мочи ниже 5,75 ураты переходят в менее растворимую форму (кетоформу) и кристаллизуются в почечных канальцах. Возрастает образование уратных камней при закислении мочи, которое возникает по различным причинам, в том числе и из-за избыточного питания мясопродуктами, которые содержат большое количество нуклеиновых кислот, амино кислот и неорганических кислот, что делает такую пищу "кислой" и усугубляет процесс.
Синдром Леша-Нихана
Болезнь Л е ша-Н и хана – это полное врожденное отсутствие активности гипоксантин-гуанин-фосфорибозил-трансферазы, фермента, отвечающего за реутилизацию пуриновых оснований. Признак рецессивный и сцеплен с Х-хромосомой. Впервые его описали в 1964 г в США студент-медик Майкл Леш и педиатр Уильям Нихан.
Дети рождаются клинически нормальными, только к 4-6 месяцу обнаруживаются отклонения в развитии, а именно – отставание физического развития (с трудом держит голову), повышенная возбудимость, рвота, периодическое повышение температуры. Выделение мочевой кислоты можно обнаружить еще раньше по оранжевой окраске пеленок. К концу первого года жизни симптомы нарастают, развивается нарушение координации движений, хореоатетоз, корковый паралич, спазм мышц ног. Наиболее характерный признак заболевания проявляется на 2-3-м году жизни – аутоагрессия или самокалечение – неодолимое стремление детей кусать себе губы, язык, суставы пальцев на руках и ногах.
Билет 57
В хранении, передаче и преобразовании генетической информации центральное место занимают нуклеиновые кислоты. Решающим фактором при этом является способность нуклеиновых, оснований к специфическому (комплементарному) спариванию (см. с. 90). (Эти процессы более детально рассматриваются в следующих разделах.)








