Для биосинтеза белка (второго этапа трансляции) требуются; мРНК как генетическая матрица, программа которой определяет порядок чередования аминокислот в белке; аминоацил-тРНК (для чтения «текста» мРНК и Как источник аминокислот при сборке белка); рибосомы как молекулярные машины для последовательного соединения аминокислот в полипептидную цепь в соответствии с программой мРНК; ГТФ как источник энергии при синтезе белка в рибосомах; белковые «факторы*, помогающие на разных фазах сборки белка в рибосомах, и, наконец, некоторые ионы как кофакторы (Mg1*, К+ и др.).
Механизм синтеза белка на рибосомах
Синтез белка, или собственно трансляцию, принято разделять на три фазы: инициация (начало), элонгация (удлинение полипепТндной цепи) и терминация (окончание).
Инициация. Начало трансляции — наиболее медленный процесс. В нерабочем состоянии субчастицы рибосом разомкнуты. мРНК, поступившая из ядра в цитоплазму, связывается с малой субчастицей на поверхности, обращенной к большой субчастнце. Причем точка присоединения к субчастнце расположена рядом с 5'-концом РНК, так как «чтение» программы РНК всегда идет в направлении 5'-*-3'. В пределах субчастицы умещаются только два кодона мРНК. Первым кодовом мРНК у 5'-конца является АУГ или ГУГ, Эти кодойы называются инициирующими, так как именно с них всегда начинается трансляция в рибосомах. Этим кодонам соответствует антикодон метионил-тРНК. У эукарнотов имеются две разные метионил-тРНК. Одна из них всегда участвует в инициации, а другая используется в процессе элонгации. У прокариотов синтез белка начинается с формилметиоиил-тРНК, где NH2-rpynna заблокирована формильной группой,
Кроме того, в инициации участвует как минимум три белковых фактора инициации (F|, Fa, F3), которые не являются составными компонентами рибосом, и ГТФ. Белковые факторы инициации облегчают связывание мРНК с малой субчастицей и ГТФ. К этому первичному комплексу (факторы инициации — малая субчастица — мРНК — ГТФ) присоединяется большая субчастица, т. е. происходит смыкание субчастиц рибосом, после чего факторы инициации удаляются из рибосом. Необходимая для смыкания субчастиц энергия получается за счет гидролиза ГТФ. Образовавшийся инициаторный комплекс (мРНК, рибосома и метионил-тРНК) готов к элонгации. Причем метионил-тРНК своим антнкодоном специфически спаривается с кодоном АУГ мРНК, т. е. как бы «подвешивается» на водородных связях к мРНК. а акцепторный конец, где находится аминокислота, прикрепляется к большой субчастице рибосом.
Элонгация. Рассмотрим, как происходит удлинение полипептида на одну аминокислоту (рис, 68), Синтез полипептида всегда начинается от N-конца и заканчивается С-концом. Наращивание полипептида на одну аминокислоту осуществляется в три шага:
1) связывание аминоацил-тРНК;
2) транспептидация (или перенос пептида);
3) транслокацня (или перемещение мРНК на один триплет).
Первый шаг. В рибосомах слева находится тРНК. которая антикодоном связана с кодоном мРНК, а акцепторный конец «связан» с растущим пептидом
Этот пептид, входящий в пептндил-тРНК. связан с П-участком, который в виде белковой ниши находится на большой субчастице. В момент первого шага второй кодон мРНК свободен. С ним спаривается своим антикодоном поступающая в рибосомы аминоацил-тРНК- Амино-ацнльный конец этой тРНК связывается с А-участ'ком большой субчастицы рибосомы. На этом первый шаг, т. е. связывание, заканчивается. На Связывание тратится энергия фосфатной связи ГТФ.
Второй шаг — транспептидация — совершается таким образом, что происходит переброс пептидила с левой тРНК на аминогруппу аминоацил-тРНК- ПрН этом образуется пептидная связь. Катализируют образование пептидной связи белки рибосом, обладающие пептидилтрансферазной активностью.
Третий шаг состоит в размыкании субчастиц рибосом, на что расходуется энергия одной молекулы ГТФ. Пептидил-тРНК, уже несущая трипепт'ид, перемешается из А-участка вместе с мРНК. с которой
она спарена, в П-участок на один триплет и выталкивает свободную тРНК нз рибосом. Для синтеза одной пептидной связи (или удлинения полипептида на одну аминокислоту) затрачивается энергия двух молекул ГТФ.
Помогают элонгации так называемые белковые факторы элонгации. Элонгация продолжается до тех пор, пока' весь текст мРНК не будет прочитан.
Термннация — окончание трансляции — зависит от присутствия в мРНК терминирующих кодонов, или «стоп-сигналов» (УАА, УГА, УАГ) и белковых факторов терминации. С терминирующими кодонами не может связаться ни одна тРНК, так как нет тРНК с соответствующими антикодокамн. Возможно, что белковые факторы терминацнк освобождают синтезированную полипептидную цепь (рис. 69).
В клетке мРНК в синтезе белка использует не одну, а несколько рибосом. Такой работающий комплекс мРНК с несколькими (от 4 до 20) рибосомами называется полирибосомой. Благодаря образованию полирибосом нет нужды в большом числе копий мРНК. В то же время синтез белка протекает быстрее, чем при использовании только одной рибосомы. За секунду полипептидная цепь удлиняется на одну аминокислоту, а в интенсивную фазу роста клеток скорость синтеза нарастает до 20 аминокислот в секунду. Послеотделения мРНК от рибосомы она тут же гидролизуется цитоплазматическими рнбонуклеазами. Поэтому для биосинтеза тех же белков необходимо вновь создавать мРНК-
Под генетическим, или аминокислотным, кодом понимают соответствие кодо-нон (кодовых слов) определенным аминокислотам. Генетический код— своеобразный словарь, переводящий текст, записанный с помощью четырех нуклеотидов, в белковый текст, записанный с помощью 20 аминокислот. Генетический код имеет следующие свойства.
1. Триплетность — каждой аминокислоте соответствует тройка нуклеотидов. Легко подсчитать, что существуют 4а=б4 кодона. Из них 61 является смысловым н 3 — бессмысленными (терминирующими).
2. Неперекрываемость — каждый из триплетов генетического текста независим друг от друга.
- 3. Вырожденность,—отдельные аминокислоты имеют несколько кодонов. Об этом говорит простое сравнение: на 20 аминокислот приходится 61 смысловой кодон, т, е. в Среднем каждой аминокислоте соответствует около 3 кодонов. Причина вырожденности кода состоит в том, что главную смысловую нагрузку несут два первых нуклеотида в триплете, а третий не так важен. Отсюда правило вырожденности кода: если два кодона имеют два одинаковых первых нуклеотида, а их третьи нуклеотиды принадлежат к одному классу (пуриновому или пиримидиновому), то они кодируют одну и ту же аминокислоту.
Однако из этого идеального правила есть два исключения. Это кодон АУА, который должен соответствовать не изолейцину, а метионину и кодон УГА, который является терминирующим, тогда как должен соответствовать Триптофану, Вырожденность кода имеет, очевидно, приспособительное значение.
4. Специфичность — каждой аминокислоте соответствуют только определенные кодоны, которые не могут использоваться для другой аминокислоты.
5. Колинеарность — соответствие линейной последовательности кодонов мРНК и аминокислот в белке.
6. Универсальность — все перечисленные выше свойства генетического кода характерны для всех жнвых организмов. В последнее время принцип универсальности кода поколеблен в связи с тем, что в митохондриях собственный генетический код отличается от известного ранее. В нем кодов УГА соответствует триптофану, а АУА — метионииу, как того требует правило вырожденности кода. Возможно, в начале эволюции у всех простейших организмов был такой же код, как и у митохондрий, а затем он претерпел небольшие отклонения.
Генетический код
способ сохранения наследственной информации в виде последовательности нуклеотидов вмолекулах нуклеиновых кислот.
Билет 62
Белки определяют жизнедеятельность клетки. Поэтому клетка должна тонко регулировать не синтез белков вообще, а необходимого в данный момент ассортимента белков.
Белки, которые синтезируются с постоянной скоростью, называются конститутивными, а синтезирующиеся с резко изменяющейся в зависимости от разных условий скоростью — адаптивными или индуцибельными. Конститутивные белки (в том числе и ферменты) содержатся в клетках примерно в постоянных количествах независимо от того, есть ли в них потребность. Количество молекул индуцибельных (адаптивных) белков варьирует в больших пределах. Очевидно, что- синтез конститутивных белков не регулируется, а индуцибельных, напротив, подвержен тонкой регулировке.
Если регуляция на уровне фермента может изменять только функциональные возможности самого фермента (интенсивна по своей природе), то регуляция синтеза белков (переноса генетической информации) изменяет количество молекул данных белков (или ферментов) и является экстенсивной.
Стимуляция биосинтеза белков, сопровождающаяся увеличением их количества, называется индукцией, а подавление синтеза белков — репрессией. Очевидно, в клетках имеются вещества, сигнализирующие о состоянии метаболизма внутри клетки или в организме. Это позволяет включать или выключать синтез белков. Такими веществами у прокариотов могут быть поступающие в клетку питательные вещества, метаболиты и некоторые внутриклеточные регуляторы (тупа циклических нуклеотидов). У многоклеточных, особенно сложноорганизованных, помимо автономных внутриклеточных регуляторов значительное место занимают внеклеточные регуляторы синтеза белков, которые подчиняют деятельность генетического аппарата биосинтеза белков конкретной клетки ткани или органа задачам целого организма.
4. Препараты, влияющие на синтез белка
Препараты, влияющие на синтез белка, широко используются в практике. Индукторы применяются с целью стимуляции синтеза белка в поврежденных или ослабленных длительным бездействием (атрофичных) органах. Этот эффект индукторов облегчает восстановление функций клеток пораженного органа.
Ингибиторы синтеза белка применяются в противоположных целях: для подавления деления и роста клеток.
Препараты, усиливающие синтез белка. Препараты этой группы являются индукторами синтеза белка и относятся к так называемым анаболическим средствам. Анаболические средства бывают гормональные и негормональные. Наиболее обширна группа препаратов гормональной природы. Среди них наиболее выраженной способностью к индукции синтеза белка (действуя на уровне транскрипции) обладают анаболические стероиды (метандростено-лон, феноболин и самый активный ретаболил), являющиеся производными мужских половых гормонов (андрогенов) и применяющиеся только с целью стимуляции синтеза белка в организме. Выраженной анаболической активностью обладает инсулин, причем этот белковый гормон, очевидно, активирует синтез белка на уровне трансляции.
Ингибиторы синтеза белка — более обширная группа препаратов, используемая при биохимических исследованиях и в практической медицине. Все ингибиторы синтеза белка можно разделить на ингибиторы: а) транскрипции,' б) процессинга и транспорта РНК, в) трансляции. Хотя некоторые препараты действуют на несколько этапов переноса генетической информации
Ингибиторы транскрипции по1 механизму делятся на три группы: ингибиторы ДНК-зависимых РНК-полимераз, блокирующие ДНК-матрицу и искажающие информацию синтезируемой РНК.
В качестве примера препаратов первой группы можно назвать: а-амани-тин (яд бледной поганки), избирательно ингибирующий РНК-полнмеразу Ш (ответственную за транскрипцию мРНК); антибиотики рифамицины, блокирующие ядрышковую РНК-полнмеразу I (отвечает за транскрипцию рРНК) и обратную транскриптазу. а-Аманитин используется при биохимических исследованиях, а рифамицины—как антибактериальные препараты " в медицинской практике.
Ко второй группе относятся вещества, связывающиеся нековалентно с матрицей ДНК и мешающие работе РНК-полимеразы. Например, актиноми-цин D (используется в биохимических исследованиях), а также антибиотики оливомицин, дактиномицин и растительные алкалоиды винбластин и вин-кристин, которые применяются в медицине как противоопухолевые препараты.
К третьей группе можно отнести, например, 5-фтороурацил, включающийся в мРНК вместо природного нуклеотида и приводящий в негодность синтезируемую матрицу РНК-Ингибиторы процессинга и транспорта мРНК- Потенциальными ингибиторами синтеза белка на этом этапе могут быть ингибиторы внутриядерных РНКаз, РНК-лигаз, осуществляющих различные фазы созревания мРНК- Препятствует присоединению полиаденилового фрагмента к мРНК кордицепин (3-дезоксиаденозин), который можно назвать ингибитором транспорта мРНК, поскольку полнадениловый фрагмент облегчает транспорт ее из ядра в цитоплазму.
Ингибиторы трансляции (т. е. синтеза белка в рибосомах). В качестве примера можно назвать антибиотики, применяемые как антибактериальные препараты.
Хлоромфеникол действует на бактериальные 70S рибосомы и рибосомы митохондрий и хлоропластов эукариотов (на 80S рибосомы он не влияет). Хлорамфеникол связывается с 50S субчастицей рибосом и блокирует пепти-дилтрансферазную реакцию, вызывая преждевременный обрыв синтезируемой полипептидной цепи.
Линкомицин близок к действию хлорамфеникол а на 80S рибосомы.
Эритромицин (и другие антибиотики макролиды) ингибирует транслокацию пептидил-тРНК из участка А в П-участок 50S субъединнцы бактериальных рибосом (т. е. блокирует третий шаг элонгации трансляции).
Тетрациклины более избирательно влияют на 70S, чем на 80S рибосомы. Блокируют связывание мРНК и аминоацил-тРНК с малой субчастицей рибосом, т. е. фазу инициации и элонгации' синтеза белка в рибосомах.
Стрептомицин влияет на 70S рибосомы бактерий и не оказывает действия на 80S рибосомы. Специфически связывается с белком малой субчастицы и нарушает правильное считывание мРНК. Синтез белка при этом прекращается или образуется дефектный белок, не способный функционировать.
В лабораторных исследованиях применяется циклогексимнд (актидион)^ действующий исключительно на 80S рибосомы эукариотов. Он связывается с большей субчастицей рибосом и тормозит транслокацию. В высоких концентрациях блокирует 'РНК-полимеразу I, т. е. действует на транскрипцию.
Билет 63
Липидами называются органические вещества биологической природы, нерастворимые в воде и растворимые в неполярных растворителях, в таких, как хлороформ, эфир или бензол.
Триацилглицериды
Триацилглицериды – это сложные эфиры глицерина и высших жирных кислот.
Общая формула:

Простые триацилглицериды содержат остатки одинаковых, смешанные – разных жирных кислот. Названия триацилглицеридов строятся на основе названий ацильных остатков, входящих в их состав жирных кислот. Смешанные триацилглицериды могут содержать хиральный атом углерода в положении 2 и иметь энантиомеры
Триацилглицериды – малополярные, не растворимые в воде вещества, так как их молекулы не содержат сильнополярных или заряженных групп. Триацилглицериды, содержащие преимущественно остатки ненасыщенных кислот, при обычных условиях являются жидкостями, насыщенных кислот – твердыми веществами. Основная биологическая функция триацилглицеридов – запасные вещества животных и растений.
Фосфолипиды
Фосфолипиды – общее название липидов, содержащих остаток фосфорной кислоты. Фосфолипиды – основные липидные компоненты клеточных мембран.
Фосфоглицериды
Основные структурные компоненты, составляющие молекулы фосфоглицеридов, – это глицерин, жирные кислоты, фосфорная кислота, аминоспирты (этаноламин или холин) или аминокислота серин. Их рассматривают как производные L-глицеро-3-фосфата, в котором спиртовые группы этерифицированы жирными кислотами, а остаток фосфорной кислоты образует сложноэфирную связь с аминоспиртом.
Главный липидный компонент клеточных мембран. Они сопутствуют жирам в пище и служат источником фосфорной кислоты, необходимый для жизни человека.
Фосфолипиды участвуют в транспорте жиров, жирных кислот и холестерина. Фосфолипиды замедляют синтез коллагена и повышают активность коллагеназы (фермента, разрушающего коллаген).
Билет 64
Переваривание жиров включает 5 этапов
1. Эмульгирование жиров пищи – необходимо для того, чтобы ферменты ЖКТ смогли начать работу.
2. Гидролиз триацилглицеролов, фосфолипидов и эфиров ХС под влиянием ферментов ЖКТ.
3. Образование мицелл из продуктов переваривания (жирных кислот, МАГ, холестерола).
4. Всасывание образованных мицелл в эпителий кишечника.
5. Ресинтез триацилглицеролов, фосфолипидов и эфиров ХС в энтероцитах.
Под влиянием перистальтики ЖКТ и составных компонентов желчи пищевой жир эмульгируется. Образующиеся лизофосфолипиды также являются хорошим поверхностно-активным веществом, поэтому они способствуют эмульгированию пищевых жиров и образованию мицелл. Гидролиз эфиров ХС осуществляет холестерол-эстераза панкреатического сока.
В панкреатическом соке также имеется активируемая трипсином фосфолипаза А2, отщепляющая жирную кислоту от С2. Обнаружена активность фосфолипазы С и лизофосфолипазы.
В результате воздействия на эмульгированные жиры ферментов панкреатического и кишечного соков образуются 2-моноацилглицерол ы, жирные кислоты и свободный холестерол, формирующие структуры мицеллярного типа (размер около 5 нм). Свободный глицерол всасывается прямо в кровь.
Синтез желчных кислот идет в эндоплазматическом ретикулуме при участии цитохрома Р450, кислорода, НАДФН и аскорбиновой кислоты. 75% холестерина, образуемого в печени, участвует в синтезе желчных кислот.
В печени синтезируются первичные желчные кислоты – холевая (гидроксилирована по С3, С7, С12) и хенодезоксихолевая (гидроксилирована по С3, С7), затем они образуют конъюгаты с глицином – гликопроизводные и с таурином – тауропроизводные, в соотношении 3:1 соответственно.
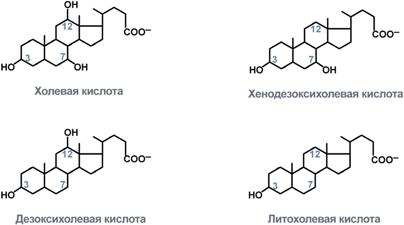
После расщепления полимерных липидных молекул полученные мономеры всасываются в верхнем отделе тонкого кишечника
Желчные кислоты частично также могут попадать в клетки и далее в кровь воротной вены, однако большая их часть остается в химусе и достигает подвздошной кишки, где всасывается при помощи активного транспорта.
Ресинтез липидов – это синтез липидов в стенке кишечника из поступающих сюда экзогенных жиров, иногда могут использоваться и эндогенные жирные кислоты. Основная задача этого процесса – связать поступившие с пищей средне- и длинноцепочечные жирные кислоты со спиртом – глицеролом или холестеролом. Это ликвидирует их детергентное действие на мембраны и позволит переносить по крови в ткани.
В кишечнике и желчи желчные кислоты находятся в виде анионов (желчные соли). По структуре желчные кислоты — амфипэтические молекулы, так как имеют гидрофобное кольцо и гидрофильные группы — гидроксильиую и карбоксильную. Поэтому они обладают выраженными поверхностно-активными свойствами, способствуя эмульгированию липидов.в кишечнике. Желчные кислоты необходимы для нормального переваривания и всасывания липидов в кишечнике. Они помогают также всасыванию в пищеварительном тракте жирорастворимых внтаминов
Билет 65
Гидролиз триацилглицеринов в тканях осуществляется тканевой триацимлицеринлипазой, которая гидролизует их на глицерин и свободные жирные кислоты. Кислая липаза содержится в лизосомах, щелочная — в микросомах, нейтральная—в цитоплазме. Характерным свойством тканевой липазы является чувствительность к гормонам, которые, активируя аденилатциклазу, вызывают переход неактивной липазы тканей в активную путем фосфорилирования с помощью протеинкиназы, Механизм этот сходен с активированием фосфорилазы В. Под действием липазы происходит мобилизация триацилглицеринов. Этот процесс называется также тканевым липолизом.
Фосфоглицериды клеточных мембран гидролизуютсн с помощью фосфолипаз А|, А2, С и D, локализованных преимущественно в лизосомах. Продуктами гидролиза фосфоглицеридов являются глицерин,'жирные кислоты, азотистые спирты, неорганический фосфат.
Окисление глицерина
Обмен глицерина тесно связан с гликолизом, в который вовлекаются метаболиты глицерина по следующей схеме:

Сначала глицерин при участии глицеролфосфокиназы превращается в альфа-глицеролфосфат. Последний под действием НАД-зависимой а-глииеролфосфат-дегидрогеназы превращается в дигидроксиацетонфосфат, который, являясь обычным метаболитом гликолиза, включается в гликолиз и превращается его ферментами до лактата в анаэробных условиях илн до СОа и НаО в аэробных. Превращение одной молекулы глицерина дает одну молекулу АТФ в анаэробных условиях и 19 молекул АТФ в аэробных. Глицерин — хороший энергетический субстрат и используется в этих целях практически всеми органами и тканями.
Эти процессы используются в качестве использования энергии.
Билет 66
Окисление жирных кислот (β-окисление)
β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С1 и С2 исходной жирной кислоты.

Элементарная схема β-окисления
Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом:
Пальмитоил-SКоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН
Этапы окисления жирных кислот
1. Активация жирной кислоты

2. Ацил-S-КоА не способен проходить через митохондриальную мембрану, поэтому существует способ его переноса в комплексе с витаминоподобным веществом карнитином. На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I.
3. После связывания с карнитином жирная кислота переносится через мембрану транслоказой. Здесь на внутренней стороне мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-S-КоА который вступает на путь β-окисления.
4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА. К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.
Значение жирных кислот как энергетических субстратов для разных органов и тканей. Не все ткани с одинаковой интенсивностью используют жирные кислоты и промежуточные продукты их окисления — кетоновые тела — как энергетические субстраты. Активно используют жирные кислоты сердце, а также почки, скелетные мышцы (при длительной работе). В этих же органах сгорают кетоновые тела, которые служат источником энергии. В нервной ткани доля использования жирных кислот и кетоновых тел для обеспечения энергетических нужд незначительна.
Билет 67
Биосинтез жирных кислот наиболее активно происходит в цитозоле клеток печени, кишечника, жировой ткани в состоянии покоя или после еды.
Условно можно выделить 4 этапа биосинтеза:
1. Образование ацетил-SКоА из глюкозы, других моносахаров или кетогенных аминокислот.
2. Перенос ацетил-SКоА из митохондрий в цитозоль:
- может быть в комплексе с карнитином, подобно тому как переносятся внутрь митохондрии высшие жирные кислоты, но здесь транспорт идет в другом направлении,
- обычно в составе лимонной кислоты, образующейся в первой реакции ЦТК.
3. Образование малонил-SКоА из ацетил-SКоА.
Карбоксилирование ацетил-SКоА катализируется ацетил-SКоА-карбоксилазой, мульферментным комплексом из трех ферментов.

Образование малонил-SКоА из ацетил-SКоА
4. Синтез пальмитиновой кислоты.
Осуществляется мультиферментным комплексом " синтаза жирных кислот " (синоним пальмитатсинтаза) в состав которого входит 6 ферментов и ацил-переносящий белок (АПБ).

Билет 68
Синтез триацилглнцеринов происходит при депонировании липидов в жировой ткани или в других тканях организма. Этот процесс локализуется в гиалоплазме клеток.
Непосредственно для синтеза триацилглнцеринов используется 3-глицерофосфат. а не свободный глицерин, и ацил-КоА, а не свободная жирная кислота, 3-глицерофосфат образуется путем фосфорилирования поступающего в ткани глицерина или при восстановлении промежуточного продукта гликолиза дигидроксиацетонфосфата.
Первой стадией синтеза триацилглицеринов служит образование фосфатндной кислоты с участием глицерофосфат-ацилтрансферазы:

Далее фосфатидная Кислота подвергается действию фосфатидат-фосфатазы с образованием днацилглицернна:

На диацилглицернн с помощью диацилглицерол-ацилтрансферазы переносится третий ацнльный остаток:

Синтезируемый триацилглицерин накапливается в виде жировых включений в цитоплазме клеток.
Биосинтез кетоновых тел
Кетоновыми, или ацетоновыми, телами называются три вещества: ацетоацетат, п-гидроксибутират и ацетон. Они являются недоокисленнымн, промежуточными продуктами распада, главным образом жирных кислот и углеродных склетов так называемых кетогенных аминокислот (лейцин, изолейции, лизин, фенилаланин, тирозин и триптофан). Образование кетоновых тел, или кето-генез, происходит в митохондриях печени.
Гидроксиметилглутаратный цикл. На первой стадии происходит конденсация двух молекул ацетил-КоА с участием ацетил-КоА-ацетилтрансферазы:

Далее ацетоацетил-КоА конденсируется еще с одной молекулой ацетил-КоА с участием гидроксиметилглутарил-КоА-синтазы:

В-Гидрокси-В-метилглутарил-КоА расщепляется под действием гидроксиметил-глутарил-КоА-лиазы на ацетил-КоА и ацетоацетат:

Ацетил-КоА вновь используется на первой стадии и тем самым переходит в цикл
. 1. Регуляция липидного обмена
1. При ограниченном потреблении углеводов или нарушении их использования (дефиците инсулина) усиливаются мобилизация жирных кислот и их транспорт кровью в печень. Происходит снижение скорости потребления ацетил-КоА путем: вовлечения в цикл трикарбоновых кислот и синтеза жирных кислот в печени.
2. При достаточном поступлении углеводов с пищей и нормальном поступлении глюкозы в клетки (что обеспечивается инсулином) увеличивается содержание метаболитов ЦТК.
Контроль скорости мобилизации и липогенеза происходит под влиянием гормонов. Активация липолиза происходит под воздействием адреналина и норадреналина, кортикостероидов, глюкагона и гормонов гипофиза — вазопрессина, АКТГ, липотропинов. Одновременно эти гормоны ограничивают стимуляцию липогенеза инсулином, результатом является повышение содержания жирных кислот в крови.
Обратный процесс — накопление липидов в депо — стимулирует инсулин. Он также активирует липогенез, обеспечивает транспорт глюкозы в клетку и ее окисление по основному пути.
Нарушения липидного обмена
Нарушения переваривания и всасывания липидов могут быть результатом:
1) процессов в поджелудочной железе с дефицитом панкреа-
тической липазы;
2) дефицита желчи в кишечнике;
3) снижения метаболической активности слизистой оболочки
кишечника, где локализованы ферменты ресинтеза триацил-
глицеридов.
Билет 69
Схема в вопросах.
Одновременно трикарбоновых кислот цикл-метаболич. путь окисления до СО2 и Н2О аминокислот, жирных к-т и углеводов, к-рые вступают в этот цикл на разл. его стадиях (схема 2). Кроме того, образующиеся ди- и трикарбоновые к-ты м. б. исходнымисубстратами в биосинтезе мн. соед. (схема 3). Так, оксало-ацетат-субстрат в глюконеогепезе; сукцинил-КоА - проме-жут. продукт в синтезе порфиринов, ацетил-КоА-в синтезе жирных к-т, стероидов, ацетилхолина. Образующийся в цикле СО2 используется в р-цияхкарбоксилирования в синтезе жирных к-т, орнитиновом цикле и др. Участие трикарбоновых кислот цикла в биосинтезе и катаболизмемн. в-в обусловливает его важное место в обмене в-в.
Билет 70
В эндокринную систему входят специальные железы, клетки которых выделяют во внутренние среды организма, т. е. в кровь или лнмфу, химические регуляторы, получившие название гормонов В настоящее время гормонами называют вещества, образующиеся в железистых клетках, выделяющиеся в кровь или лимфу и регулирующие обмен веществ и развитие организма.
По химическому строению гормоны подразделяются на:
1) белково-пептидные (гормоны гипоталамуса, гипофиза, поджелудочной и паращитовидной желез, кальциотоннн щитовидной железы);
2) производные аминокислот (адреналин — производное аминокислот феиилаланина и тирозина; иодтиронины — производные тирозина; мелатонин - производное триптофана);
3) стероиды (половые гормоны — андрогены, эстрогены и гестагены; кортнкостероиды)
Биологическое действие гормонов проявляется через их взаимодействие с рецепторами клеток-мишеней. Для проявления биологической активности связывание гормона с рецептором должно приводить к образованию химического сигнала внутри клетки, который вызывает специфический биологический ответ, например изменение скорости синтеза ферментов и других белков или изменение их активности. Мишенью для гормона могут служить клетки одной или нескольких тканей. Воздействуя на клетку-мишень, гормон вызывает специфическую ответную реакцию. Характерный признак клетки-мишени - способность воспринимать информацию, закодированную в химической структуре гормона.
Начальный этап в действии гормона на клетку-мишень - взаимодействие гормона с рецептором клетки. Клетки-мишени отличают соответствующий гормон от множества других молекул и гормонов благодаря наличию на клетке-мишени соответствующего рецептора со специфическим центром связывания с гормоном.
Связывание гормона (первичного посредника) с рецептором приводит к изменению конформации рецептора. Это изменение улавливается другими макромолекулами, т.е. связывание гормона с рецептором приводит к сопряжению одних молекул с другими (трансдукция сигнала). Таким образом, генерируется сигнал, который регулирует клеточный ответ путём изменения активности или количества ферментов и других белков. В зависимости от способа передачи гормонального сигнала в клетках меняется скорость реакций метаболизма:
- в результате изменения активности ферментов;
- в результате изменения количества ферментов
Гормоны (первичные посредники), связываясь с рецепторами на поверхности клеточной мембраны, образуют комплекс гормон-рецептор, который трансформирует сигнал первичного посредника в изменение концентрации
Билет 71
Регуляция деятельности желез внутренней секреции осуществляется нервными и гуморальными факторами.
Регулирующее влияние ЦНС на деятельность эндокринных желез осуществляется через гипоталамус. Гипоталамус получает по афферентным путям мозга сигналы из внешней и внутренней среды. Нейросекреторные клетки гипоталамуса трансформируют афферентные нервные стимулы в гуморальные факторы, продуцируя рилизинг-гормоны. Рилизинг-гормоны избирательно регулируют функции клеток аденогипофиза. Среди рилизинг-гормонов различают либерины - стимуляторы синтеза и выделения гормонов аденогипофиза и статины - ингибиторы секреции. Они носят название соответствующих тропных гормонов: тиреолиберин, кортиколиберин, соматолиберин и т.д. В свою очередь, тропные гормоны аденогипофиза регулируют активность ряда других периферических желез внутренней секреции (кора надпочечников, щитовидная железа, гонады). Это так называемые прямые нисходящие регулирующие связи.
К настоящему времени в гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины) секреции гормонов гипофиза, а именно: кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин, пролактолиберин, меланолиберин,соматостатин, пролактостатин и меланостатин.
Установлено, что по химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми олигопептидами необычного строения.
Гипоталамические гормоны непосредственно влияют на секрецию (точнее, освобождение) «готовых» гормонов и биосинтез этихгормонов de novo.
Билет 72
Мембранно-внутриклеточный тип действия характерен для гормонов (и других внеклеточных регуляторов), которые не проникают в клетку и поэтому влияют на обмен веществ через внутриклеточный химический посредник (вторичный посредник), который и является своеобразным полномочным представителем гормона внутри клетки.
Внутриклеточные посредники — цАМФ и цГМФ, ионы Са2+ и 2', 5'-олиго (А)я-адениловый олигонуклеотид. Гормоны регулируют образование вторичных посредников в клетке, а те, в свою очередь, влияют на активность и количество разных ферментов и тем самым изменяют биохимические функции клетки.
Внеклеточные регуляторы действуют на образование циклических нуклеотидов через одну из двух сигнальных систем: аденилатциклазу или гуанилатциклазу. Первая контролирует образование цАМФ, вторая — цГМФ:
Аденилатциклаза встроена в мембрану. Как только гормон связывается с рецептором, то комплекс гормон — рецептор взаимодействует с N-белком. В результате изменяется конфигурация N-белка и происходит связывание ГТФ с этим белком, а правильнее сказать, замещение ГДФ, находящегося в неактивном N-белке, на ГТФ. Комплекс N-белок — ГТФ является аллостерическим активатором собственно аденилатциклазы (каталитической части). Активация аденилатциклазы приводит к наработке цАМФ внутри клетки из АТФ.В активном состоянии аденилатциклаза поддерживается до тех пор, пока существует комплекс гормон--рецептор, который обеспечивает образование комплекса N-белок — ГТФ, активирующего аденилатциклазу. При диссоциации комплекса гормон — рецептор прекращается его дейст








