Динамика структуры хромосом в клеточном цикле.
Митоз -тип деления клетки, при котором образуются дочерние клетки с таким же набором хромосом, как и у материнской клетки.
| Фаза
| Процессы
|
| Профаза
| 1. Хромосомы спирализуются, в результате чего становятся видимыми. 2. Каждая хромосома состоит из двух хроматид. 3. Ядерная мембрана и ядрышко разрушаются. Центриоль удваивается.
|
| Метафаза
| 4. Хромосомы располагаются по экватору клетки. Образуется веретено деления.
|
| Анафаза
| 5. Центромеры делятся, и хроматиды (дочерние хромосомы) расходятся к полюсам клетки с помощью нитей веретена деления.
|
| Телофаза
| 6. Вокруг разошедшихся хромосом образуется новая ядерная мембрана. 7. Исчезает веретено деления. Образуются две дочерние клетки.
|
Регуляторная роль компонентов хромосом заключается в «запрещении» или «разрешении» считывания информации с молекулы ДНК.
В первой половине митоза хромосомы состоят из двух хроматид. соединенных между собой в области первичной перетяжки (центромеры) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
Механизмы упаковки ДНК в метафазную хромосому
Перед делением клетки молекулы ДНК удваиваются, петли укладываются в стопки, хромосома утолщается и видна в световой микроскоп. На этом уровне упаковки каждая хромосома состоит из двух хроматид. Каждая из хроматид содержит по одной молекуле ДНК.
14. Морфологическая и функциональная характеристика хромосом. Гетеро- и эухроматин.
а) Гетерохроматин. Во время интерфазы определённые участки хромосом и целые хромосомы остаются компактными. Они образуют «глыбки» интенсивно окрашенные и, как правило, прилежащие к мембране ядра. Гетерохроматин неактивен в отношении транскрипции. Существует две формы гетерохроматина: факультативный и конститутивный
• факультативный гетерохроматин бывает гетерохроматичным только временами. Он информативен, содержит гены с которых считывается наследственная информация, когда гетерохроматин переходит в эухроматическое состояние. Образуется при спирализации одной из двух гомологичных хромосом. Типичным примером служит тельце полового хроматина, образуемого одной из двух Х-хромосом соматических клеток женских особей человека и млекопитающих. Функциональная роль факультативного гетерохроматина заключается в компенсации снижении дозы определенного гена (например, появление промежуточного признака при явлении неполного доминирования у гетерозигот Аа, влияет на экспрессивность проявления наследственных признаков в фенотип), определяет тканеспецифичность.
• структурный гетерохроматин – отличается высокоспирализованным состоянием, которое сохраняется на протяжении всего мит. цикла. Он занимает постоянные участки в гомологичных хромосомах – это фрагменты околоцентромерных, теломерных участков хромосом, Не содержит структурных генов (нетранскрибируемый); Его роль не ясна, но по видимому он выполняет опорную функцию.
б) Эухроматин (разрыхленный) - имеет менее компактную организацию, деспирализуется в конце митоза, образует слабоокрашенные нитчатые структуры содержит структурные транскрибируемые гены;
Метафазная хромосома
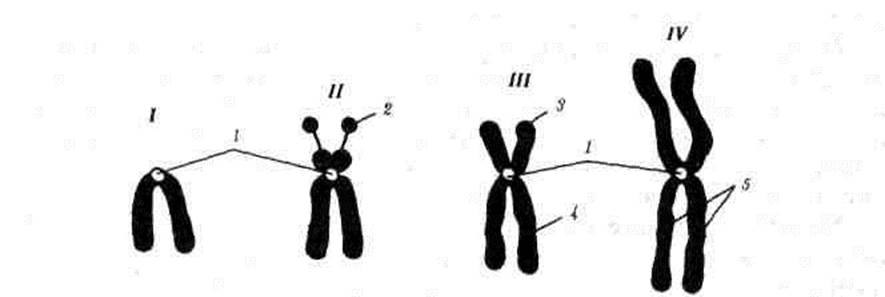 Формы хромосом
Формы хромосом
/—телоцентрическая, //—акроцентрическая, ///—субметацентрическая, IV —метацентрическая;
1-—центромера, 2 —спутник, 3 —короткое плечо «p», 4 —длинное плечо –«q », 5 — хроматиды
1. В соматических клетках диплоидный (двойной) набор хромосом – 2п4с.
2. В диплоидном наборе хромосомы парные.
3. Парные хромосомы имеют одинаковое строение и называются – г о м о л о г и ч н ы е.
4. Хромосомы из разных пар – н е г о м о л о г и ч н ы е.
5. Хромосомы, имеющие одинаковое строение в клетках особей разного пола, называются,
а у т о с о м а м и. Их обозначают арабскими цифрами (1,2,3,…). Они представлены парами гомологичный хромосом, но индивидуально различных (отцовских и материнских). Их располагают в порядке уменьшения размеров, поэтому самая большая хромосома имеет первый номер. У человека самая маленькая аутосома имеет 22 номер.
У человека в соматических клетках 22 пары - 44 аутосомы, а в половых клетках - 22 аутосомы
6. Хромосомы участвующие в определении пола, называются п о л о в ы м и или
г е т е р о х р о м о с о м а м и (гетеросомы), их обозначают латинскими буквами «Х» и «У». В соматических клетках человека две половые хромосомы, у женского пола две ХХ, у мужского ХУ.
Т.о. в соматических клетках человека 46 хромосом = 44 аутосомы две половые ХХ, или 44 аутосомы + две половые ХУ, а в половых клетках 23 хромосомы = 22 аутосомы + одна половая Х или 22 аутосомы + одна половая У.
7. В половых клетках (гаметах – сперматозоидах и яйцеклетках) содержится половинный - г а п л о и д н ы й – п набор хромосом.
15. Сцепление генов. Кроссинговер. Митотический кроссинговер. Неравный кроссинговер.
Кроссинговер – это процесс обмена гомологичными участками гомологичных хромосом(хроматид)
Митотический кроссинговер:
• Происходит рекомбинация маркеров одной группы сцепления
• Происходит на стадии 4-х нитей
• Обмениваются 2 несестринские хромосомы
• В результате обмена образуются реципрокные продукты
Неравный кроссинговер:
Кроссинговер, в результате которого образуются сестринские кроссоверные хроматиды, различающиеся поколичеству заключенного в них генетического материала; в основе Н.к. лежит неправильное спариваниедуплицированных сегментов рекомбинирующих хроматид (например, в составе кластера генов рРНК), врезультате которого в одной хроматиде остается 1 копия, а в другой - 3 копии данного сегмента
Сцепление генов:
Полное сцепление - гены, относящиеся к одной группе сцепления, всегда наследуются вместе
Неполное сцепление – между генами, относящимися к одной группе сцепления, возможна рекомбинация
Классификация генов.
Гены классифицируют на структурные и функциональные.
Структурные гены – кодируют развитие конкретных признаков белка, фермента, РНК. Функциональные гены оказывают влияние на работу структурных генов. Функциональными являются гены-модуляторы и гены-регуляторы. Гены-модуляторы – это ингибиторы, интенсификаторы, модификаторы. Они усиливают, ослабляют или изменяют работу структурных генов. Регуляторы (их больше всего) координируют активность структурных генов (сайленсеры-понижают активность генов, энхансеры– повышают активность, инсуляторы – предотвращают активность.
Протоонкогены – гены, функционирующие в эмбриогенезе, участвующие в контроле клеточного роста. Во взрослом состоянии блокируются.
Окогены – гены, продукты которых (онкобелки) вызывают злокачественные перерождения клеток (малигнизация).
Антионкогены – препятствуют действию онкобелкови деблокировке протоонкогенов.
Псевдогены – эволюционный мусор, не транскрибируются, либо с них считывается функционально неактивный белок.
Гены домашнего хозяйства(конститутивные) – функционируют во всех клетках. Независимо от их тканевой принадлежности. Это гены гистонов, тРНК. рРНК и тд. Обеспечивают процессы репликации, транскрипции, трансляции.
Гены роскоши(регулируемые) – гены, активные в клетках определенных тканей, в определенное время.
Мобильные гены – участки генома, которые могут перемещаться как внутри одной хромосомы, так и между хромосомами.
Перемещения: нереплекативная транспозиция (вырезать и вставить).
Репликативная транспозиция (копировать ДНК и вставить)
РНК-опосредованная транспозиция (копировать РНК – обратная транскрипция – ДНК и вставить)
Значения: приводит к наследственной изменчивости, вызывает мутации, изменяет активность рядом расположенных генов, горизонтальный перенос генов, используются в генной инженерии.
Прокариоты. 1)геном содержит порядка 1000 генов,локализованных в одной кольцевоймолекулеДНК нуклеоида-ядерного аппарата бактерий. 2)отсутствие интронов в генах. 3)имеют более сложные транскриптоны, содержащие группу последовательно расположенных СГ,на одном фланге которой находится Ини, а на другой Тер. Такойпрокариотический транскриптон наз-ся опероном.
Эукариоты. 1) генетич.материал имеется в ядерном и в некоторых органоидах, поэтому геном эукариот состоит из нескольких разных компонентов. Ядерный геном содержит ядерные гены-нуклеом. Митохондриальный геном содержит митохондриальные гены-хондриом. Пластидныйгеном содержит пластидные гены-пластидом. 2)экзон-интронная структура(мозаичное строение). 3)каждый транскриптон включает в себя один СГ и два РГ.
20. Генетическая инженерия. Методы, задачи, перспективы.
Генная инженерия — это метод биотехнологии, который занимается исследованиями по перестройке генотипов.
Генотип является не просто механическая сумма генов, а сложная, сложившаяся в процессе эволюции организмов система. Генная инженерия позволяет путем операций в пробирке переносить генетическую информацию из одного организма в другой. Перенос генов дает возможность преодолевать межвидовые барьеры и передавать отдельные наследственные признаки одних организмов другим.
Перестройка генотипов, при выполнении задач генной инженерии, представляет собой качественные изменения генов не связанные с видимыми в микроскопе изменениями строения хромосом. Изменения генов прежде всего связано с преобразованием химической структуры ДНК. Информация о структуре белка, записанная в виде последовательности нуклеотидов, реализуется в виде последовательности аминокислот в синтезируемой молекуле белка. Изменение последовательности нуклеотидов в хромосомной ДНК, выпадение одних и включение других нуклеотидов меняют состав образующихся на ДНК молекулы РНК, а это, в свою очередь, обуславливает новую последовательность аминокислот при синтезе. В результате в клетке начинает синтезироваться новый белок, что приводит к появлению у организма новых свойств. Сущность методов генной инженерии заключается в том, что в генотип организма встраиваются или исключаются из него отдельные гены или группы генов. В результате встраивания в генотип ранее отсутствующего гена можно заставить клетку синтезировать белки, которые ранее она не синтезировала.
Наиболее распространенным методом генной инженерии является метод получения рекомбинантных, т.е. содержащих чужеродный ген, плазмид. Плазмиды представляют собой кольцевые двухцепочные молекулы ДНК, состоящие из нескольких тысяч пар нуклеотидов. Этот процесс состоит из нескольких этапов.
ü Рестрикция — разрезание ДНК, например, человека на фрагменты.
ü Лигирование — фрагмент с нужным геном включают в плазмиды и сшивают их.
ü Трансформация — введение рекомбинантных плазмид в бактериальные клетки. Трансформированные бактерии при этом приобретают определенные свойства. Каждая из трансформированных бактерий размножается и образует колонию из многих тысяч потомков — клон.
ü Скрининг — отбор среди клонов трансформированных бактерий тех, которые плазмиды, несущие нужный ген человека.
Весь этот процесс называется клонированием. С помощью клонирования можно получить более миллиона копий любого фрагмента ДНК человека или другого организма. Если клонированный фрагмент кодирует белок, то экспериментально можно изучить механизм, регулирующий транскрипцию этого гена, а также наработать этот белок в нужном количестве. Кроме того, клонированный фрагмент ДНК одного организма можно ввести в клетки другого организма.
Овца Долли:
Генетическая информация для процесса клонирования была взята из взрослых дифференцированных (соматических) клеток, а не из половых (гамет) или стволовых. Самого исходного животного (прототипа) на момент клонирования уже не существовало. А часть его клеток, необходимая для эксперимента, была своевременно заморожена и хранилась в жидком азоте, чтобы сохранить и передать генетический материал.
Перспективы
Активно ведутся исследования по разработке вакцин для профилактики и лечения гепатитов, СПИДа и ряда других заболеваний, а также конъюгированных вакцин нового поколения против наиболее социально значимых инфекций. Полимер-субъединичные вакцины нового поколения состоят из высокоочищенных протективных антигенов различной природы и носителя – иммуностимулятора полиоксидония, обеспечивающего повышенный уровень специфического иммунного ответа. Прививки против подавляющего большинства известных инфекций Россия могла бы обеспечить на базе собственного иммунологического производства. Полностью отсутствует только производство вакцины против краснухи.
Виды изменчивости:
• Ненаследственная (фенотипическая, определенная)
• Наследственная (генотипическая, неопределенная)
• Онтогенетическая
Формы изменчивости
1. Ненаследственная модификационная(фенотипическая)
Изменение условий среды, в результате чего организм изменяется в пределах нормы реакции, заданной генотипом.
2 Наследственная (генотипическая) Мутационная
Влияние внешних и внутренних мутагенных факторов, в результате чего происходит изменение в генах и хромосомах.
Комбинативная
Возникает стихийно в рамках популяции при скрещивании, когда у потомков появляются новые комбинации генов.
3 механизма:
Два из них связаны с мейозом. При кроссинговере аллельный состав хромосом половых клеток потомка изменяется в сравнении с таковы гамет родителей. В связи с неизвестным характером расхождения отцовских и материнских хромосом представлены одновременно хромосомы унаследованные от обоих родителей, третий механизм связан с оплодотворением. При случайном подборе гамет создается разнообразие сочетаний хромосом.
Модификационная изменчивость отражает изменение фенотипа под воздействием факторов внешней среды (усиление и развитие мышечной и костной массы у спортсменов, увеличение эритропоэза в условиях высокогорья и крайнего севера).
Мутационная изменчивость играет роль главного поставщика наследственных изменений. Именно она является первичным материалом всех эволюционных преобразований.
Одним из распространенных видов мутаций, имеющий важное значение в эволюции растений, является полиплоидия.
Хромосомные мутации также играют важную эволюционную роль. Прежде всего необходимо указать на удвоение генов в одной хромосоме. Именно благодаря удвоениям генов в процессе эволюции накапливается генетический материал. Нарастание сложности организации живого в ходе исторического развития в значительной степени опиралось на увеличение количества генетического материала.
Генные мутации – наиболее частый тип мутаций. Мутации отдельных генов происходят редко. Большинство мутаций рецессивные, доминантные мутации возникают намного реже. Доминантные и рецессивные мутации ведут себя в популяциях по-разному. Доминантные мутации, даже если они находятся в гетерозиготном состоянии, проявляются в фенотипах особей уже первого поколения и подвергаются действию естественного отбора. Рецессивные же мутации проявляются в фенотипе только в гомозиготном состоянии.
Рецессивная мутация, прежде чем она проявится в фенотипе гомозигот, должна накопиться в значительном количестве в популяции. Эту мысль первым высказал выдающийся советский генетик Четвериков.
Он писал, что популяция, подобно губке, впитывает рецессивные мутации, оставаясь при этом фенотипически однородной. Существование такого скрытого резерва наследственной изменчивости создает возможность для эволюционных преобразований популяций под воздействием естественного отбора.
Комбинативная изменчивость – это следствие перекреста гомологичных хромосом, их случайного расхождения в мейозе и случайного сочетания гамет при оплодотворении. Комбинативная изменчивость ведет к появлению бесконечно большого разнообразия генотипов и фенотипов. Она служит неиссякаемым источником наследственного разнообразия видов и основой для естественного отбора.
Громадное генотипическое и, следовательно, фенотипическое разнообразие в природных популяциях является тем исходным эволюционным материалом, с которым оперирует естественный отбор.
Свойства модификаций:
1. Прямопропорциональность – сила воздействия фактора прямо прямо пропорциональна появлению модификация.
2. Адаптивность – большинство адаптаций имеют приспособительный характер
3. Определенность – факторы действуют только на определенный признак
4. Групповой характер – меняется целая группа организмов
5. Обратимость
6. Ненаследуемость
Адаптивные модификации – реакции клеток и организма на изменения условий среды, которые неоднократно действовали на организм в ходе эволюции
Неадаптивные модификации – не имеют приспособительного значения и вызываются экстремальными воздействиями (морфозы, тераты, фенокопии.)
Механизмы модификационной изменчивости:
1) Окружающая среда как причина модификаций. Модификационная изменчивость — это результат не изменений генотипа, а его реакции на условия окружающей среды. При модификационнойизменчивости наследственный материал не изменяется, — изменяется проявление генов. (пр-р: выработка меланина)
2) Норма реакции. Предел проявления модификационной изменчивости организма при неизменном генотипе — норма реакции. Норма реакции обусловлена генотипом и различается у разных особей данного вида. Фактически норма реакции — спектр возможных уровней экспрессии генов, из которого выбирается уровень экспрессии, наиболее подходящий для данных условий окружающей среды. Норма реакции имеет предел для каждого вида (пр-р: размеры листьев растений, жирность молока у коров)
Генетическая обусловленность нормы реакции:
1. Полигенная детерминация признака и реакции организма
2. Плейотропность генов - это зависимость нескольких признаков от одного гена
3. Зависимость проявления гена от условий среды
4. Гетерозиготность организма
5. Взаимодействие генов, которое происходит на уровне генных продуктов (субъединиц белковых молекул)
Механизмы:
1) рекомбинация генов при кроссинговере
2) свободное расхождение гомологичных хромосом в анафазе мейоза-1 и хроматид в анафазе мейоза 2.
3) случайное сочетание гамет при оплодотворении
4) взаимодействие аллельных и неаллельных генов
Примеры: У цветка ночная красавица есть ген красного цвета лепестков А, и ген белого цвета а. Организм Аа имеет розовый цвет лепестков. Таким образом, у ночной красавицы нет гена розового цвета, розовый цвет возникает при сочетании (комбинации) красного и белого гена.
У человека есть наследственное заболевание серповидноклеточная анемия. АА – норма, аа – смерть, Аа – СКА. При СКА человек не может переносить повышенных физических нагрузок, при этом он не болеет малярией, т.е. возбудитель малярии малярийный плазмодий не может питаться неправильным гемоглобином. Такой признак полезен в экваториальном поясе; для него нет гена, он возникает при сочетании генов А и а.
Значение: - для организма может быть:
а) адаптивной, обеспечивает приспособление к условиям среды. Например, малярия и серповидноклеточная анемия.
б) нейтральной, не влияет на приспособленность,но при определенных изменениях среды обитания могут оказаться полезными для организмов.
- для эволюции: повышает генетическое разнообразие внутри популяции вида благодаря возникновению новых сочетаний генов в генотипе.
26. Онтогенетическая изменчивость у человека, ее механизмы и примеры.
Онтогенетическая изменчивость — изменчивость, в процессе индивидуального развития которой происходят закономерные изменения морфологических, физиологических, биохимических признаков.
Причины:
1. Понижение активности генов, приводящих к нарушению метаболизма.
2. Различие времени экспрессии генов
Особенности:
1. имеют общие закономерности для всех особей данного вида
2. подвержена влиянию внешней среды
3. необратимый характер
Онтогенетическая изменчивость на примере гемоглобина:
Виды:
1) Эмбриональный (HbE) – формируется в желточном мешке, 6-8 неделя внутриутробного развития
2) Фетальный (HbF) – плодный, форм-ся в печени и селезенке
3) Дефинитивный (HbA) – взрослый, форм-ся с момента рождения
HbF начинает синтезироваться с 12-14 недели развития плода (с момента формирования плаценты), заменяя эмбриональный гемоглобин. Его первичная структура отличается от первичной структуры HbA в тридцати девяти позициях (последовательность β-цепей по сравнению с таковой у γ-цепей). Эти отличия лежат в основе отличий физико-химических свойств HbA от HbF. Фетальный гемоглобин является устойчивым к денатурирующему воздействию щёлочи. Это отличительное свойство легло в основу метода количественного определения фетального гемоглобина. Кроме того, фетальный гемоглобин в большей степени способен превращаться в метгемоглобин, имеет специфический спектр поглощения в ультрафиолетовой части спектра. К моменту рождения на долю фетального гемоглобина приходится 80-85 % от
общего количества. Синтез фетального гемоглобина в течение первого года жизни замедляется, заменяясь гемоглобином взрослого типа. К трём годам его количество соответствует содержанию HbF у взрослого человека и составляет 1-1,5 %.
Механизмы:
1) функциональное различие генов в процессе онтогенеза (пр-р: гемоглобин)
2) повышение и снижение активности генов (пр-р: синтез коллагена)
Внутрихромосомные
1. дупликации- удвоение участка хромосомы Dp(13;13)
2. делеции - выпадение срединного участка хромосомы Del (5р)
3. З. дефишенси - выпадение концевого участка Df21q)
4. Инверсии: перицентрические - вокруг центромерыinv 5(21 p31q); парацентрические - около центромеры
Межхромосомные транслокации
1. Реципрокные транслокации — это сбалансированные хромосомные перестройки, при которых весь генетический материал сохраняется, а изменения касаются только расположения генов в хромосомах. (например: Синдром Дауна tr 21\13(15); синдром Прадера-Вилли 46ХУ tr 15\на любую
2. Нереципрокная (несбалансированная)
3. Роберсоновская - объединение 2х центрических хромосом в одну, или разделение метацентрической на 2 (! изменяется число хромосом)
Хромосомный перестройки
1. Изохромосома - возникает когда центромераделится не продольно а поперечно. В результате одно из плеч теряется, а 2ое удваивается (46Xi(Xq) - синдром ломкой хромосомы)
2. Кольцевые хромосомы - разрывы в обоих плечах. Ацентричеческие фрагменты теряются, а центральная часть замыкается в кольцо.
СОВЕТУЮ ПОЧИТАТЬ ЕЩЕ НА СТРАНИЦЕ 128-133 в Ярыгине
Полиплоидия и гетероплоидия, их механизмы. Явление нерасхождения хромосом как причина возникновения хромосомных болезней. Примеры.
Геномные мутации — это мутации, которые приводят к добавлению либо утрате одной, нескольких или полного гаплоидного набора хромосом.
1) Полиплоидия (эуплоидия) – увеличение числа кратного гаплоидному набору
• Сбалансированная – 4n, 6n, 8n.
• Несбалансированная – 3n, 5n, 7n.
o Автоплоидия – увеличивается за счет хромосомодного вида.
o Аллоплоидия – за счет хромосом разных видов.
2) Гетероплоидия (анэуплоидия) – изменение числа хромосом некратно гаплоидному набору (2n+1)
Примеры:
Нарушение числа аутосом
• С-м Патау 47XX13+
• С-м Эдвардса 47XУ18+
• С-м Дауна 47XX21+
Спонтанные мутации.
Мутации, помимо качественных свойств, характеризует и способ возникновения. Спонтанные (случайные) – мутации, возникающие при нормальных условиях жизни. Спонтанный процесс зависит от внешних и внутренних факторов (биологические, химические, физические). Спонтанные мутации возникают у человека в соматических и генеративных тканях. Метод определения спонтанных мутаций основан на том, что у детей появляется доминантный признак, хотя у его родителей он отсутствует.
Индуцированные мутации.
Индуцированный мутагенез – это искусственное получение мутаций с помощью мутагенов различной природы. Впервые способность ионизирующих излучений вызывать мутации была обнаружена Г.А. Надсоном и Г.С. Филлиповым.Затем, проводя обширные исследования, была установлена радиобиологическая зависимость мутаций. В 1927 году американским ученым Джозефом Мюллером было доказано, что частота мутаций увеличивается с увеличением дозы воздействия.
Мутагены — химические и физические факторы, вызывающие наследственные изменения — мутации.
Мутаген - фактор, вызывающий мутации.
По природе возникновения мутагены классифицируют на физические, химические и биологические:
Физические мутагены.
-ионизирующее излучение;
-радиоактивный распад;
-ультрафиолетовое излучение;
-моделированное радиоизлучение и электромагнитные поля;
-чрезмерно высокая или низкая температура.
Химические мутагены.
-окислители и восстановители (нитраты, нитриты, активные формы кислорода);
-алкилирующие агенты (например, иодацетамид);
-пестициды (например гербициды, фунгициды);
-некоторые пищевые добавки (например, ароматические углеводороды, цикламаты);
-продукты переработки нефти;
-органические растворители;
-лекарственные препараты (например, цитостатики, препараты ртути, иммунодепрессанты).
К химическим мутагенам условно можно отнести и ряд вирусов (мутагенным фактором вирусов являются их нуклеиновые кислоты — ДНК или РНК).
Биологические мутагены.
-специфические последовательности ДНК — транспозоны;
-некоторые вирусы (вирус кори, краснухи, гриппа);
-продукты обмена веществ (продукты окисления липидов);
-антигены некоторых микроорганизмов.
Генеалогический метод.
Метод основан на прослеживании какого-либо признака в ряде поколений с указанием родственных связей (составление родословной).
Сбор сведений начинается от пробанда.
Пробанд - лицо, родословную которого необходимо составить. Братья и сестры пробанда называются сибсы.
Метод включает два этапа:
1. Сбор сведений о семье.
2. Генеалогический анализ.
Для построения родословной применяются специальные символы.
Методы позволяют установить:
1.установить факт передачи признака по наследству
2.определить тип и характер наследования
3.дать прогноз (вероятность) рождения больного ребенка
4.характер сцепления генов
5.пенетрантость
При аутосомно-доминантном наследовании ген проявляется в гетерозиготном состоянии у лиц обоих полов; сразу в первом поколении; большое количество больных, как по вертикали, так и по горизонтали. По такому типу наследуются веснушки, брахидактилия, катаракта, хрупкость костей, хондродистрофическаякарликовость, полидактилия.
При аутосомно-рецессивном наследовании мутационный ген проявляется только в гомозиготном состоянии у лиц обоего пола. Как правило, у здоровых родителей (ген в гетерозиготном состоянии) рождаются больные дети. Признак проявляется не в каждом поколении. Так наследуются признаки: Леворукость, рыжие волосы, голубые глаза, миопатия, сахарный диабет, фенилкетонурия.
При Х-сцепленном доминантном наследовании болеют лица обоего пола, чаще встречается у женщин. Так наследуются признаки: пигментный дерматоз, кератоз (потеря волосяного покрова), пузырчатость стоп ног, коричневая эмаль зубов.
При Х-сцепленном рецессивном наследовании больны в основном лица мужского пола. В семье больна половина (50%) мальчиков 50% девочек гетерозиготны по мутантному гену. Так наследуется гемофилия А, мышечная дистрофия Дюшена, дальтонизм.
При У-сцепленном наследовании больны только мужчины. Такие признаки называются голандрические: синдактилия, гипертрихоз.
Близнецовый метод.
Это один из наиболее ранних методов изучения генетики человека, однако, он не утратил своего значения и в настоящее время.
Близнецовый метод был введен Ф.Гамильтоном, который выделил среди близнецов две группы:
•однояйцевые (монозиготные)
•двуяйцевые (дизиготные)
Монозиготные близнецы при нормальном эмбриональном развитии всегда одного пола. Дизиготные близнецы рождаются чаще (2/3 общего количества двоен), они развиваются из двух одновременно созревших и оплодотворенных яйцеклеток. Такие близнецы могут быть и однополые и разнополые. С генетической точки зрения они сходны как обычные сибсы, но у них большая общность факторов среды во внутриутробном(пренатальном) и частично в постнатальном периодах.
Если изучаемый признак проявляется у обоих близнецов пары, их называют конкордантными. Конкордантность – это процент сходства по изучаемому признаку. Отсутствие признака у одного из близнецов – дискордантность. Близнецовый метод используется в генетике человека для того, чтобы оценить степень влияния наследственности и среды на развитие какого-либо нормального или патологического признака. Для оценки роли наследственности в развитии того или иного признака производят расчет по формуле:
Н = (% сходства ОБ - % сходства ДБ) / (100 - % сходства ДБ)
где:
Н- коэффицент наследственности
ОБ – однояйцевые близнецы
ДБ – двуяйцевые близнецы
•При Н = 1 признак полностью определяется наследственным компонентом
•При Н = 0 признак определяется влиянием среды
•При Н = близкий к 0,5 признак определяется примерно одинаковым влиянием наследственности и среды на формирование признака
Этапы:(лекция)
1.взятие материала (забор клетки)
2.культивирование клеток (митогенные факторы, ФГА-фитогемоглютинин (вроде так хз)
3. накопление метафазных клеток
4.гипотонизация клеток
5.окрашивание препарата
6.составление идиограммы и ее анализ
Инвазивные
1. Амниоцентез - забор околоплодной жидкости и находящихся в ней слущенных клеток амниона и плода на 15-18 неделе беременности для цитогенетических, молекулярно-генетических, биохимических методов.
2. Биопсия хориона и плацентобиопсия - получение кусочков ворсин хориона или кусочков плаценты в период с 8 по 16 неделю для цитогенетического, молекулярно-генетического и биохимических методов в исследования.
3. Кородоцентез - взятие крови из пуповины в период с 12 по 22 недели для цитогенетического, молекулярно-генетического и биохимических методов исследования.
4. Биопсия тканей плода (кожи, мышц)- биопсия кожи, мышц для гистологического анализа.
5. Фетоскопия (! редко)- визуальный осмотр плода при введении зонда в амниотическую полость на 18-23 неделе.
По объёму
Большие
Средние
Маленькие
Идеальная популяция - абстрактная, бесконечно большая, только панмиксия, не действуют элементарные эволюционные факторы (в природе не существует) Экологические характеристики
Размер ареала - зависит от радиуса индивидуальной активности
Численность (ниже минимальной численности популяция исчезает по зколого-генетическим причинам
Плотность - отношение кол-ва особей на единицу площади
Возрастная структура - зависит от уровня рождаемости и смертности, скоростей смены поколений и продолжительности жизни
Половая структура - соотношение полов в разных возрастных группах
Соотношение полов
первичное - на момент зачатия 1:1
вторичное - на момент рождения - Ж 100:М 106
третичная
16-18 лет 100:100
50 лет 100:85
85 лет 100:50
Генетические характеристики:
Генофонд (А.С. Серебровский 1928 - генофонд - совокупность генотипов всех особей популяции)
Тип размножения:
• Панмиксия - свободной скрещивание (более устойчивое)
• Инбридинг - близкородственное скрещивание, приводящее к обеднению генофонда и гомогенизации популяции
Скорость обновления
Генетический полиморфизм - присутствие в генофонде нескольких аллелей отдельных генов
Генетическое единство - общая реакция популяции на изменения условий окружающей среды
Генетический гомеостаз - способность популяции поддерживать единство своего генетического состава
Генетический груз - насыщенность генофонда мутациями, снижающими приспособленность популяции
Популяция людей - группа людей, занимающая одну территорию длительное время и свободно вступающая в брак Демографические характеристики
1.Размер
большие (200 тыс)
средние Ф
малые
демы (до 4 тыс, частота внутрнгрупповыкбраков до 80%о)
изоляты (до 1,5 тыс, частота близкородственных браков до 90%)
2. Уровень рождаемости и смертности
3. Возрастной н половой состав
4. Уклад жизни (городское или сельское)
5. Социально-экономическое состояние_
Дрейф генов (генетико-автоматические процессы) - случайное колебание частот аллелей в популяциине обусловленные действием естественного отбора. Заметен в малых популяциях - в изолятах (1931-1932 Райт и Дубинин) изоляция как движущая сила эволюции.
Динамика структуры хромосом в клеточном цикле.
Митоз -тип деления клетки, при котором образуются дочерние клетки с таким же набором хромосом, как и у материнской клетки.
| Фаза
| Процессы
|
| Профаза
| 1. Хромосомы спирализуются, в результате чего становятся видимыми. 2. Каждая хромосома состоит из двух хроматид. 3. Ядерная мембрана и ядрышко разрушаются. Центриоль удваивается.
|
| Метафаза
| 4. Хромосомы располагаются по экватору клетки. Образуется веретено деления.
|
| Анафаза
| 5. Центромеры делятся, и хроматиды (дочерние хромосомы) расходятся к полюсам клетки с помощью нитей веретена деления.
|
| Телофаза
| 6. Вокруг разошедшихся хромосом образуется новая ядерная мембрана. 7. Исчезает веретено деления. Образуются две дочерние клетки.
|
Регуляторная роль компонентов хромосом заключается в «запрещении» или «разрешении» считывания информации с молекулы ДНК.
В первой половине митоза хромосомы состоят из двух хроматид. соединенных между собой в области первичной перетяжки (центромеры) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.






