

Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Топ:
Определение места расположения распределительного центра: Фирма реализует продукцию на рынках сбыта и имеет постоянных поставщиков в разных регионах. Увеличение объема продаж...
Генеалогическое древо Султанов Османской империи: Османские правители, вначале, будучи еще бейлербеями Анатолии, женились на дочерях византийских императоров...
Интересное:
Средства для ингаляционного наркоза: Наркоз наступает в результате вдыхания (ингаляции) средств, которое осуществляют или с помощью маски...
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Подходы к решению темы фильма: Существует три основных типа исторического фильма, имеющих между собой много общего...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Перед делением клетки молекулы ДНК удваиваются, петли укладываются в стопки, хромосома утолщается и видна в световой микроскоп. На этом уровне упаковки каждая хромосома состоит из двух хроматид. Каждая из хроматид содержит по одной молекуле ДНК.
14. Морфологическая и функциональная характеристика хромосом. Гетеро- и эухроматин.
а) Гетерохроматин. Во время интерфазы определённые участки хромосом и целые хромосомы остаются компактными. Они образуют «глыбки» интенсивно окрашенные и, как правило, прилежащие к мембране ядра. Гетерохроматин неактивен в отношении транскрипции. Существует две формы гетерохроматина: факультативный и конститутивный
• факультативный гетерохроматин бывает гетерохроматичным только временами. Он информативен, содержит гены с которых считывается наследственная информация, когда гетерохроматин переходит в эухроматическое состояние. Образуется при спирализации одной из двух гомологичных хромосом. Типичным примером служит тельце полового хроматина, образуемого одной из двух Х-хромосом соматических клеток женских особей человека и млекопитающих. Функциональная роль факультативного гетерохроматина заключается в компенсации снижении дозы определенного гена (например, появление промежуточного признака при явлении неполного доминирования у гетерозигот Аа, влияет на экспрессивность проявления наследственных признаков в фенотип), определяет тканеспецифичность.
• структурный гетерохроматин – отличается высокоспирализованным состоянием, которое сохраняется на протяжении всего мит. цикла. Он занимает постоянные участки в гомологичных хромосомах – это фрагменты околоцентромерных, теломерных участков хромосом, Не содержит структурных генов (нетранскрибируемый); Его роль не ясна, но по видимому он выполняет опорную функцию.
|
|
б) Эухроматин (разрыхленный) - имеет менее компактную организацию, деспирализуется в конце митоза, образует слабоокрашенные нитчатые структуры содержит структурные транскрибируемые гены;
МОРФОЛОГИЯ МЕТАФАЗНЫХ МИТОТИЧЕСКИХ ХРОМОСОМ.
Для изучения индивидуального набора хромосом (кариотипа) особое значение имеют митотические метафазные хромосомы т. к. хромосомы на этой стадии максимально спирализованы и видны как отдельные морфологические структуры. Благодаря спирализации достигается плотная упаковка наследственного материала, что важно для перемещения хромосом в процессе митоза
1. Хромосома состоит из двух половинок - хроматид, каждая хроматида состоит из биспирали ДНК. Хромосомы обозначают – «п», ДНК – «с», т.о. хромосома – П 2С
2. На теле хромосомы есть первичная перетяжка – центромера или кинетохор
Центромера делит тело хромосомы на плечи. В зависимости от расположения центромеры различают хромосомы по форме: метацентрические (равноплечие), субметацентрически (неравноплечие имеют длинное плечо –«q» и короткое - «p»), акроцентрические (палочковидные), телоцентрические (выражено неравноплечие);
4. Некоторые хромосомы имеют вторичную перетяжку, которая отделяет
небольшой участок – спутник хромосомы. Хромосома, имеющая спутник называется спутничной. У человека спутничные хромосомы относятся к аутосомам и имеют номер – 13, 14, 15, 21, 22
5. В области вторичных перетяжек некоторых хромосом располагаются я д р ы ш к о в ы е о р г а н и з а т о р ы. Они содержат гены, кодирующие рРНК и служат местом образования я д р ы ш к а.
6. На концах плеч хромосом расположены т е л о м е р ы. Они препятствуют склеиванию хромосом, возможно содержат гены, отвечающие запродолжительность жизни.
Метафазная хромосома
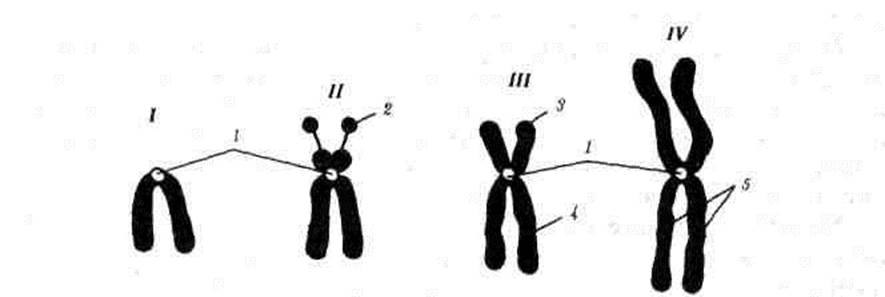 Формы хромосом
Формы хромосом
/—телоцентрическая, //—акроцентрическая, ///—субметацентрическая, IV —метацентрическая;
|
|
1-—центромера, 2 —спутник, 3 —короткое плечо «p», 4 —длинное плечо –«q », 5 — хроматиды
1. В соматических клетках диплоидный (двойной) набор хромосом – 2п4с.
2. В диплоидном наборе хромосомы парные.
3. Парные хромосомы имеют одинаковое строение и называются – г о м о л о г и ч н ы е.
4. Хромосомы из разных пар – н е г о м о л о г и ч н ы е.
5. Хромосомы, имеющие одинаковое строение в клетках особей разного пола, называются,
а у т о с о м а м и. Их обозначают арабскими цифрами (1,2,3,…). Они представлены парами гомологичный хромосом, но индивидуально различных (отцовских и материнских). Их располагают в порядке уменьшения размеров, поэтому самая большая хромосома имеет первый номер. У человека самая маленькая аутосома имеет 22 номер.
У человека в соматических клетках 22 пары - 44 аутосомы, а в половых клетках - 22 аутосомы
6. Хромосомы участвующие в определении пола, называются п о л о в ы м и или
г е т е р о х р о м о с о м а м и (гетеросомы), их обозначают латинскими буквами «Х» и «У». В соматических клетках человека две половые хромосомы, у женского пола две ХХ, у мужского ХУ.
Т.о. в соматических клетках человека 46 хромосом = 44 аутосомы две половые ХХ, или 44 аутосомы + две половые ХУ, а в половых клетках 23 хромосомы = 22 аутосомы + одна половая Х или 22 аутосомы + одна половая У.
7. В половых клетках (гаметах – сперматозоидах и яйцеклетках) содержится половинный - г а п л о и д н ы й – п набор хромосом.
15. Сцепление генов. Кроссинговер. Митотический кроссинговер. Неравный кроссинговер.
Кроссинговер – это процесс обмена гомологичными участками гомологичных хромосом(хроматид)
Митотический кроссинговер:
• Происходит рекомбинация маркеров одной группы сцепления
• Происходит на стадии 4-х нитей
• Обмениваются 2 несестринские хромосомы
• В результате обмена образуются реципрокные продукты
Неравный кроссинговер:
Кроссинговер, в результате которого образуются сестринские кроссоверные хроматиды, различающиеся поколичеству заключенного в них генетического материала; в основе Н.к. лежит неправильное спариваниедуплицированных сегментов рекомбинирующих хроматид (например, в составе кластера генов рРНК), врезультате которого в одной хроматиде остается 1 копия, а в другой - 3 копии данного сегмента
|
|
Сцепление генов:
Полное сцепление - гены, относящиеся к одной группе сцепления, всегда наследуются вместе
Неполное сцепление – между генами, относящимися к одной группе сцепления, возможна рекомбинация
|
|
|

Типы сооружений для обработки осадков: Септиками называются сооружения, в которых одновременно происходят осветление сточной жидкости...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...

История развития пистолетов-пулеметов: Предпосылкой для возникновения пистолетов-пулеметов послужила давняя тенденция тяготения винтовок...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!