Физиологические механизмы долговременной памяти не могут базироваться на циркуляции импульсов или изменениях электрофизиологических характеристик отдельных нейронов. При различных воздействиях на организм (гипоксия, наркоз, охлаждение, электрошок) разрушаются кольцевые реверберационные связи, и снижается возбудимость нейронов. Но при этом огромное количество информации сохраняется в долговременной памяти в неизменном виде.
Долговpеменную память нельзя связать с увеличением количества нейронов, поскольку они не способны к воспроизведению. Однако их аксоны могут медленно удлиняться, если впеpеди есть свободное пpостpанство, и обpазовывать дополнительные синапсы. Одновpеменно с этим могут дополнительно ветвиться дендpиты, обpазуя дендpитные шипики и маленькие волосковые выступы. Благодаpя таким стpуктуpным изменениям пpоисходит обpазование новых синапсов или pеактивный синаптогенез. Чем богаче жизненный опыт pастущих животных, тем большим у них становится объем тех или иных отделов коpы, пpичем пpиpост связан с клетками нейpоглии.
Согласно анатомической теории запоминание и хранение информации осуществляется за счет образования новых терминальных волокон, изменения их размеров, развития шипикового аппарата на дендритах нейронов. Такие структурные изменения предполагают увеличение темпа синтеза белка в нейронах. Действительно, давно и точно установлено, что блокирование процессов транскрипции в нейроне, нарушение синтеза РНК и белка полностью блокирует консолидацию и формирование энграммы памяти. Следует обратить внимание на один важный факт: в подавляющем большинстве соматических клеток экспрессируется не более 7% генома, а в клетках ЦНС, особенно в нейронах коры головного мозга до 25%, а в некоторых отделах до 38%. Это свидетельствует о том, что возможности для синтеза белков у нейронов больше, а белки разнообразнее. Естественно, никаких новых белков, кроме закодированных в структурах ДНК, синтезироваться не может. Что же это за белки? Сразу следует сказать о том, что далеко не для всех синтезируемых в процессе консолидации белков установлена их функциональная роль. Есть, например, белок, получивший название S-100, этот и ряд других белков пока не «получили» своего назначения, хотя их концентрация значительно увеличивается в процессе формирования энграммы памяти. Предполагается их участие в фосфорилировании других белков, модулировании рецепторного ответа. Остановимся на тех белках, значение которых более понятно.
Во-первых, это структурные белки нейронов, синтез которых приводит к увеличению количества и размеров шипиков на нейронах, а значит и установлению более прочных межнейроных связей. В экспериментах на животных показано, что в коре головного мозга детенышей, выращенных в сенсорно обедненной среде, нейроны менее ветвистые, у них меньше дендритов, меньше шипиков, да и толщина коры в целом значительно уступает коре сверстников, выращенных в нормальных условиях.
Во-вторых, это рецепторные белки и белки ионных каналов. Вернемся еще раз к глутаматным рецепторам. Оказывается, они принимают участие не только в механизмах кратковременной памяти – установлено, что при обучении активируется синтез этих рецепторов и увеличивает их число на постсинаптической мембране.
В-третьих, это регуляторные белки. Например, нейрогормон АКТГ приводит к фосфориллированию ряда мембранных белков (В-50) и стойкому снижению мембранного потенциала покоя, определяя тем самым длительное повышение возбудимости нейронов. Запоминание улучшается в присутствии вазопрессина, ангиотензина, а окситоцин, наоборот, нарушает долговременную память.
Наконец, это медиаторные белки, синтез которых всегда увеличивается в процессе запоминания. Например, установлено, что повышение концентрации норадреналина в структурах ЦНС ускоряет обучение в случае использования отрицательного подкрепления, а серотонин – в случае положительного. Кроме того, есть еще один механизм, повышающий эффективность работы синапсов – установлено, что в нейронах различных областей ЦНС не только увеличивается количество ацетилхолина, но и возрастает активность холинэстеразы. Следовательно, повышается функциональная лабильность синапса и частота проведения импульса.
Велика, как выяснилось, роль ионов кальция в механизмах долговременной памяти. Ионы кальция не только деполяризуют мембрану, что отражается в процессе посттетанической потенциации, но и изменяют метаболизм нейронов. Накопление ионов кальция в цитоплазме нейронов приводит к двум важным последствиям. 1. Ионы кальция, активируя специфическую протеинкиназу, увеличивают чувствительность мембранных рецепторов. 2. Под действием ионов кальция происходит фосфориллирование регуляторных белков хроматина, следовательно, увеличивается доступ к ДНК, и вновь процессы синтеза белка.
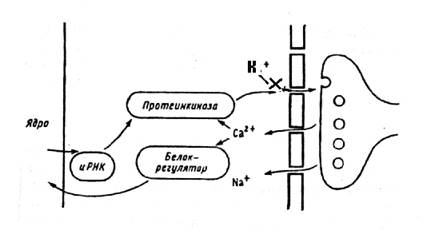
Рисунок 7
На рисунке представлены механизмы участия ионов кальция в долговременной потенциации – блокирование калиевых каналов, и долговременной памяти – активация синтеза РНК, синтез протеинкиназ и изменение количества мембранных рецепторов.
Существует глиальная теория долговременной памяти. Эта теория основывается на изменениях глиальных клеток, которые окружают нейроны и могут синтезировать особые вещества, облегчающие синаптическую передачу или повышающие возбудимость соответствующих нейронов. Нейpоглия обеспечивает необходимое питание нейpонов и напpавляет pост их аксонов. Кpоме того, она обpазует миелиновое покpытие неpвных волокон, что повышает эффективность передачи возбуждения по наиболее часто используемым путям. Установлено, что изменения электpических потенциалов, возникающие в нейpонах, pождают потенциалы и в клетках глии.
Кроме того, известно, что глия способствует более быстрому восстановлению мембранного потенциала нейрона, следовательно, увеличивается его функциональная лабильность. Как видим, и эта гипотеза требует синтеза новых структурных компонентов. Действительно, экспериментально доказано, что обогащение среды, в которой происходит рост животного, увеличивает и скорость миелинизации нервных волокон, и толщину миелинового слоя. Таким образом, существует зависимость между обогащением жизненного опыта и моpфологическими изменениями в наиболее активно используемых стpуктуpах мозга.
Интересна гипотеза об иммунохимическом механизме долговременной памяти. Автор этой гипотезы И.П. Ашмарин считает, что при длительной реверберации импульса в нейронной ловушке на синаптических мембранах синтезируются антигены, а в глиальных клетках антитела к ним. Следовательно, появляется дополнительный механизм контакта между нейронами и глиальными клетками.








