Глюкоза

 2АТФ
2АТФ
I ЭТАП: 2НАДН → дых. цепь → 6АТФ
2ПВК
На I этапе синтезируются 2 молекулы АТФ в расчёте на одну молекулу глюкозы. Если гликолиз завершается образованием двух молекул лактата (восстанавливаются две молекулы пирувата с помощью ЛДГ и двух молекул НАДН), то в аэробных условиях пируват не восстанавливается, а подвергается декарбоксилированию, т.е. две молекулы НАДН «экономятся». Реакции гликолиза протекают в цитоплазме клетки, и образовавшийся здесь НАДН не может передавать водород непосредственно в дыхательную цепь, поскольку митохондриальная мембрана для этого кофермента непроницаема. Перенос водорода с цитозольного НАДН внутрь митохондрии осуществляется специальными механизмами – челночными. Один из них значимый – малат – аспартатный.
Суть его работы заключается в том, что НАДН в цитозоле клетки восстанавливает оксалоацетат, который при этом превращается в малат, затем малат проникает внутрь митохондрии, где отдаёт восстановленный кофермент в дыхательную цепь.
Таким образом, за счёт двух молекул НАДН, образовавшихся в цитоплазме клетки на стадии гликолитической оксидоредукции (окисление трёхуглеродных молекул в пируват, который восстанавливается в лактат), дополнительно синтезируется 3 · 2 = 6 молекул АТФ (дыхательная цепь, окисляющая НАДН, имеет 3 пункта фосфорилирования – это I, II, IV комплексы = 3 молекулы АТФ и за счёт 2-х молекул НАДН → 3 · 2 = 6АТФ). Всего на I этапе получаем 2 АТФ (синтезированы в реакциях субстратного фосфорилирования (6-я, 9-я реакции) до стадии образования пирувата + 6АТФ = 8АТФ).
2 ПВК
 II ЭТАП: 2НАДН → дых. цепь → 6АТФ
II ЭТАП: 2НАДН → дых. цепь → 6АТФ
2CH3CO – S – КоА
На 2-м этапе при окислительном декарбоксилировании двух молекул пирувата в митохондриях получаем 6 молекул АТФ за счёт 2-х молекул НАДН, образующихся под действием пируватдегидрогеназы.
Пируватдегидрогеназный комплекс участвует в окислительном декарбоксилировании пирувата с образованием ацетил - КоА.
Комплекс включает три фермента и пять коферментов:
1) пируватдегидрогеназа с коферментом ТПФ
2) дигидролипоилтрансацетилаза с коферментом амидом липоевой кислоты и коферментом А.
3) дигидролипоилдегидрогеназа, имеющая кофакторы ФАД и НАД.
 2CH3CO – S – КоА
2CH3CO – S – КоА
 2 АТФ
2 АТФ
 III ЭТАП: 6НАДН → дых. цепь → 18АТФ
III ЭТАП: 6НАДН → дых. цепь → 18АТФ
 2ФАДН2 → дых. цепь → 4АТФ
2ФАДН2 → дых. цепь → 4АТФ
6CO2 + H2O
Пируватдегидрогеназный комплекс участвует в окислительном декарбоксилировании пирувата с образованием ацетил - КоА.
Комплекс включает три фермента и пять коферментов:
1) пируватдегидрогеназа с коферментом ТПФ
2) дигидролипоилтрансацетилаза с коферментом амидом липоевой кислоты и коферментом А.
3)дигидролипоилдегидрогеназа, имеющая кофакторы ФАД и НАД.
На 3-м этапе при окислении двух молекул ацетил-КоА за счёт водороддонорной и собственно энергетической функции цикла Кребса получаем 12 · 2 = 24 АТФ.
В сумме на всех 3-х этапах аэробного окисления 1 моля глюкозы получаем 38 молей АТФ. Как видно, в энергетическом отношении этот процесс много выгоднее гликолиза.
Полная энергия распада глюкозы составляет 2880 КДж/моль. Свободная энергия гидролиза высокоэнергетической связи АТФ равна 50 КДж/моль. Для синтеза АТФ при окислении глюкозы используется 38 · 50 = 1900 КДж, что составляет 65% от всей энергии распада глюкозы. Это максимально возможная эффективность использования энергии глюкозы.
Пентозофосфатный путь в метаболизме глюкозы.
В результате исследований было установлено, что в печени, почках и форменных элементах крови, особенно во время усиленного обмена углеводов, образуются пентозо-фосфорные эфиры.
Образование пентоз из глюкозы происходит путём декарбоксилирования и называется апотомическим путем. Последовательность реакций пентозофосфатного пути можно разделить на 2 части: окислительную и неокислительную, а также двух фаз: аэробной и анаэробной.
В результате окислительного пути (аэробная фаза) образуется 2 молекулы НАДФН2, которые не окисляются дыхательной цепью, а служат источником водорода и электронов при синтезах жирных кислот, холестерина, стероидных гормонов, ацетоновых тел и др. соединений. В результате окислительной части образуется фосфорилированная пентоза – рибулозо-5-фосфат.
В результате неокислительного пути (анаэробная фаза) образуется рибозо-5-фосфат. Этот углевод и его производные используются для синтеза РНК, ДНК, АТФ, КоА, НАД и ФАД.
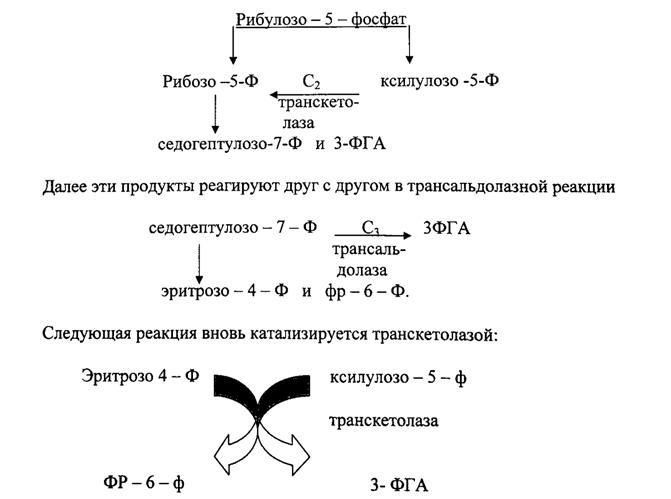
Неокислительная часть пути включает 2 типа реакций: транскетолазную и трансальдолазную. Коферментом транскетолазы является тиаминпирофосфат.
Транскетолаза переносит С2-фрагменты, трансальдолаза – С3-фрагменты.
У млекопитающих активность пентозофосфатного пути высока в печени, жировой ткани, надпочечниках, лактирующей молочной железе.
Пентозофосфатный путь тесно связан с гликолизом. У обоих путей превращения глюкозы имеются общие метаболиты: фр-6-ф и 3-ФГА. Фр-6-ф
 гл-6-ф, а 3-ФГА может участвовать не только в гликолизе, но и в
гл-6-ф, а 3-ФГА может участвовать не только в гликолизе, но и в
фосфогексо-
изомераза
аэробном пути окисления глюкозы. От гликолиза пентозофосфатный путь существенно отличается тем, что в нём участвуют не НАД+, а НАДФ+; в апотомическом пути образуется CO2 (чего нет в гликолизе) и он не обладает энергетической функцией.
Регуляция. Ключевыми ферментами пентозофосфатного пути являются дегидрогеназа гл-6-ф и дегидрогеназа 6-фосфоглюконата (окислительная часть). Индуктором их биосинтеза является инсулин. Эти дегидрогеназы считаются адаптивными ферментами, т.к. их активность зависит от питания – при достаточном поступлении углеводов они активируются. При диабете, голодании – ингибируются.
Лекция № 12
ТЕМА «ОБМЕН УГЛЕВОДОВ»
Содержание темы:
1. Глюконеогенез, регуляция.
2. Роль печени в углеводном обмене.
3. Регуляция углеводного обмена.
Глюконеогенез
Глюконеогенез – это синтез глюкозы из неуглеводных предшественников. У млекопитающих эту функцию выполняет в основном печень, в меньшей мере – почки и клетки слизистой оболочки кишечника. Запасов гликогена в организме достаточно для удовлетворения потребностей в глюкозе в период между приемами пищи. При углеводном или полном голодании, а также при длительной физической работе концентрация глюкозы в крови поддерживается за счет глюконеогенеза. В этот процесс могут быть вовлечены вещества, которые способны превратиться в пируват или любой другой метаболит глюконеогенеза.
Использование первичных субстратов в глюконеогенезе происходит при различных физиологических состояниях. Так, в условиях голодания часть тканевых белков распадается до аминокислот, которые затем используются в глюконеогенезе. При распаде жиров образуется глицерин, который через диоксиацетонфосфат включается в глюконеогенез. Лактат, образующийся при интенсивной физической работе в мышцах, в печени превращается в глюкозу. Следовательно, физиологическая роль глюконеогенеза из лактата, аминокислот и глицерина различна.
Глюконеогенез в основном протекает по тому же пути, что и гликолиз, но в обратном направлении. Однако имеется очень важная особенность, обусловленная тем, что 3 реакции в гликолизе, катализируемые киназами: гесокиназой, фосфофруктокиназой и пируваткиназой, необратимы и на этих стадиях глюконеогенеза отличаются от реакций гликолиза. Ключевые ферменты глюконеогенеза:
1. Пируваткарбоксилаза;
2. Фосфоенолпируваткарбоксикиназа;
3. гл-6-фосфотаза;
4. фр-6-бисфосфотаза.
Превращение пирувата в фосфоенолпируват осуществляется при участии двух ферментов – пируваткарбоксилазы (а) и карбоксикиназы фосфоенолпирувата (б):
пируваткарбоксилаза
 Пируват + АТФ + СО2 + Н2О ЩУК + АДФ + Н3РО4 (а),
Пируват + АТФ + СО2 + Н2О ЩУК + АДФ + Н3РО4 (а),
фосфоенолпируват-
 ЩУК + ГТФ карбоксикиназа фосфоенолпируват + СО2 + ГДФ (б).
ЩУК + ГТФ карбоксикиназа фосфоенолпируват + СО2 + ГДФ (б).
Две другие необратимые реакции катализируются фосфатазой фруктозо-1,6-бисфосфата и фосфатазой глюкозо-6-фосфата:
Фруктозо-1,6-бисфосфат +Н2О → фруктозо-6-фосфат + Н3РО4,
Глюкозо-6-фосфат + Н2О → глюкоза + Н3РО4.
На каждую молекулу лактата при глюконеогенезе расходуется три молекулы АТФ (точнее, две АТФ и одна ГТФ); поскольку для образования глюкозы необходимо 2 молекулы лактата, суммарный процесс глюконеогенеза описывается так:
2 лактат + 6 АТФ + 6 Н2О → глюкоза + 6 АДФ + 6 Н3РО4.
Образовавшаяся глюкоза может вновь поступать в мышцы и там превращаться в молочную кислоту.
Сравним реакцию глюконеогенеза с суммарной реакцией гликолиза:
Глюкоза + 2 АДФ + 2 Н3РО4 → 2 лактат + 2 АТФ + 2 Н2О.
Из этого сопоставления следует, что в результате действия глюкозо-лактатного цикла Кори работающие мышцы добывают 2 АТФ за счет расходования 6 АТФ в печени.











