

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Определение места расположения распределительного центра: Фирма реализует продукцию на рынках сбыта и имеет постоянных поставщиков в разных регионах. Увеличение объема продаж...
Интересное:
Влияние предпринимательской среды на эффективное функционирование предприятия: Предпринимательская среда – это совокупность внешних и внутренних факторов, оказывающих влияние на функционирование фирмы...
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Берегоукрепление оползневых склонов: На прибрежных склонах основной причиной развития оползневых процессов является подмыв водами рек естественных склонов...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Рассмотренные выше процессы, способные изменить генетический состав популяции и вызвать эволюционные изменения, в естественных условиях чрезвычайно редко действуют в чистом виде. На протяжении одного поколения популяция подвергается действию мутационного процесса, дрейфа генов, естественного отбора и миграции. Дифференцировать изменения по природе вызывающих их процессов - трудная и часто неразрешимая задача. Один из подходов к решению этой проблемы заключается в определении условий, при которых решающим оказывается один, или, в крайнем случае, два эволюционных фактора.
В работах Айала [1], Гранта [6] приведены соотношения, позволяющие оценить совместное действие дрейфа генов и таких факторов, как мутации, отбор и миграции. Интенсивность каждого из них можно выразить посредством коэффициентов u, s, m. По Айала дрейф генов будет основным фактором, когда произведение 4·N·xi<< 1, где xi - один из упомянутых коэффициентов, N - эффективный размер популяции. Когда величина выражения близка или больше единицы, мутации, отбор или миграция становятся решающими. Грант [6] определил условия для преимущественного действия дрейфа генов при величинах выражения 2·N·xi£1, а для их совместного действия интервал, в котором величина эффективной численности популяции 0.25xi£ N £ 0.5 xi.
Другим подходом может быть построение несложных "однолокусных" математических моделей, характеризующих изменение частоты контролируемого аллеля за одно поколение. При этом требуется допущение, что действие эволюционных факторов аддитивно. Например, при сочетании отбора и мутационного процесса общее изменение частоты аллеля равно сумме изменений частот в каждом отдельном процессе
|
|
Dqобщ = Dqотб + Dqмут (73)
Dqобщ.= pq[q(W2-W1) + p(W1- Wo)]/Ws + pu - qv, (74)
где u и v скорости прямого и обратного мутирования.
Построение графических зависимостей Dqобщ от q позволяет в ряде случаев выявить дополнительные точки равновесия или найти изменения положения для уже существующих.
Генетическая дифференциацияв процессе видообразования
Одним из проявлений процесса эволюции является внутри- и межвидовая изменчивость. Различия в фенотипических признаках основаны на существующих генотипических различиях. Последние обнаруживают себя, когда удается получить второе поколение потомков при скрещивании близкородственных видов. Однако в рамках менделевской генетики невозможно ответить на вопрос, какова степень генетических различий между популяциями одного вида или между различными видами. С другой стороны, ответив на этот вопрос, в рамках монофилетической концепции организменной эволюции можно не только установить степень родства, но и сравнить "эволюционные расстояния", отделяющие представителей двух различных видов друг от друга и от их "общего" предка.
В настоящее время существует несколько методов для оценки генетической дифференциации двух популяций. Можно упомянуть метод оценки тождественности геномов и более подробно остановиться на методе анализа электрофоретических свойств белков. Полагают, что каждому аллелю в локусе соответствует своя изоформа белка, отличающаяся от другой изоформы своей подвижностью в токопроводящем геле.
Масатоши Ней [1] предложил дифференцировать две популяции на основе электрофоретического анализа белков. В качестве характеристик генетической дифференциации предложено два параметра: генетическое сходство I как доля структурных генов, которые идентичны в обеих популяциях, и генетическое расстояние D как среднее число замен аллелей в каждом локусе.
Рассмотрим пример оценки генетической дифференциации четырех видов приматов (табл. 9) на основе анализа 19 локусов, кодирующих белки крови. В столбцах для каждого вида приведены цифровые индексы аллелей (96, 97, 98 и т.д.). В случаях, когда локус полиморфен, в скобках указаны соответствующие частоты аллелей.
|
|
Таблица 9
| Локус | Шимпанзе | Горилла | Гиббон | Бабуин |
| 1. Ak | 98 (0.20) 100 (0.80) | |||
| 2. Alb | ||||
| 3. Aph | ||||
| 4. Cer | ||||
| 5. Dia | 85 (0.67) 95 (0.33) | 100 (0.67) 108 (0.33) | 95 (0.88) 100 (0.12) | |
| 6. Est-A | ||||
| 7. Est-B | 95 (0.17) 96 (0.08) 103 (0.75) | |||
| 8. G6pd | ||||
| 9. Got | 96 (0.14) 100 (0.86) | |||
| 10. Hb | 100 (0.92) 102 (0.08) | |||
| 11. Hpt | ||||
| 12. Icd | 100 (0.94) 107 (0.06) | |||
| 13. Lap | ||||
| 14. Ldh-A | ||||
| 15. Ldh-B | ||||
| 16. Mdh | 93 (0.62) 100 (0.38) | |||
| 17. 6-Pgd | 97 (0.15) 105 (0.85) | |||
| 18. Pgm-1 | 96 (0.12) 100 (0.88) | |||
| 19. Pgm-2 |
Для расчета генетического сходствадвух популяций по одному локусу используют следующее выражение:
 , (75)
, (75)
где ai и bi - частоты аллелей i - локуса в популяциях А и В.
В качестве примера рассмотрим популяции шимпанзе и гориллы. Локус Alb (2) мономорфен и представлен одинаковым аллелем с индексом 100. Частоты этого аллеля a100 и b100 равны единице и показатель генетического сходства равен

Локус Cer (4) в каждой популяции мономорфен, но представлен различными аллелями с индексами 100 и 98. Показатель генетического сходства равен:

Локус 6-Pgd (17) в популяции шимпанзе мономорфен и представлен аллелем с индексом 97, в популяции гориллы - полиморфен (аллели с индексами 97 и 105). В этом случае показатель генетического сходства равен:

Степень генетической дифференциации двух популяций оценивают по нескольким локусам. Для этого можно воспользоваться формулой, которую предложил М.Ней [1]:
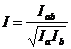 (76)
(76)
где Iab средняя сумма произведений частот одноименных аллелей:
 , (77)
, (77)
Ia средняя сумма квадратов частот в первой популяции:
 , (78)
, (78)
Ib тоже для второй популяции:
 . (79)
. (79)
В рассмотренном примере:




От показателя генетического сходства можно перейти к показателю генетического расстояния D, равного  . В нашем примере D= ‑ln0.392 = 0.936. Это означает, что за время раздельной эволюции в трех рассмотренных локусах в среднем произошло 0.936 замены или 93.6%.
. В нашем примере D= ‑ln0.392 = 0.936. Это означает, что за время раздельной эволюции в трех рассмотренных локусах в среднем произошло 0.936 замены или 93.6%.
Результаты определения генетического сходства I и генетического расстояния D между представленными выше приматами даны в табл. 10. Величины I и D показаны над и под диагональю соответственно.
|
|
Таблица 10
| Шимпанзе | Горилла | Гиббон | Бабуин | |
| Шимпанзе | Ý | 0.60000 | 0.4844 | 0.3764 |
| Горилла | 0.5108 | Ý | 0.5756 | 0.4466 |
| Гиббон | 0.7248 | 0.5523 | Ý | 0.5050 |
| Бабуин | 0.9771 | 0.8061 | 0.6833 | Ý |
Результаты анализа можно представить графически, расположив виды приматов в углах пирамиды, длины ребер которой равны генетическому расстоянию (рис.10).
 |
Рис. 10. Распределение в воображаемом пространстве видов, в соответствии с межвидовыми генетическими расстояниями.
Однако, когда число сравниваемых популяционных систем становится больше 4, графическое решение теряет смысл, так как для N видов требуется «N-1»-мерное пространство.

Рис. 11. Гипотетическое филогенетическое древо некоторых приматов
Когда изучаемые виды эволюционно связаны, их родословную определяют следующим образом. Эволюционно продвинутый вид (в нашем примере это шимпанзе) помещают на вершину. Ниже располагают вид генетически наиболее приближенный к первому (горилла). Затем определяют среднее расстояние между этими двумя видами и гиббоном (0.7248+0.5523)/2=0.6385 и между теми же двумя видами и бабуином (0.9771+0.8061)/2=0.8916. Сравнение средних величин показывает, что гиббон, по-видимому, ближе к шимпанзе и горилле, чем бабуин. Следовательно, наиболее правдоподобна схема филогении, показанная на рис. 11.
Список литературы
1. Айала Ф. Введение в популяционную и эволюционную генетику. - М.: Мир, 1984. - 230 с.
2. Реймерс И.Ф. Основные биологические понятия и термины. – М.: Просвещение, 1988.- 319 с. Грант В. Эволюционный процесс. - М.: Мир, 1991. - 488 с.
3. Кайданов Л.З. Генетика популяций. - М.: Высшая школа, 1996.- 320 с.
4. Кимура М. Молекулярная эволюция: теория нейтральности. - М.: Мир, 1985.- 394 с.
5. Северцов А.С. Основы теории эволюции. М.: Из-во МГУ, 1987.- 556 с.
6. Грант В. Эволюционный процесс. М.: Мир, 1991. 448 с.
Предметный указатель
адаптационная ценность генотипа..... 34
гальтоновские признаки........................ 5
генетико-автоматические процессы... 23
генетическая дифференциация............ 47
Генетический груз................................ 43
|
|
генетическое сходство.......................... 48
Генотипическое разнообразие............... 5
гетерозиготность..................................... 8
гетерозиготность максимальная ожидаемая 9
гетерозиготность полиморфных локусов 8
гетерозиготность теоретически ожидаемая 9
группы крови.......................................... 5
динамика генетического груза............ 44
дрейф генов........................................... 23
естественный отбор.............................. 34
инбридинг.......................................... 7, 25
Количество генотипов........................... 7
коэффициент инбридинга.................... 26
коэффициент отбора............................. 34
критерий полиморфности...................... 8
максимальная гетерозиготность популяции 13
менделевские признаки.......................... 5
миграция................................................ 31
множественные аллели.......................... 7
Мутации................................................. 28
отбор гамет............................................ 45
панмиксия.............................................. 15
панмиктическая популяция................. 14
плата за отбор.................................. 43, 44
полиморфность........................................ 8
полифакториальные генотипы............ 18
поток генов............................................ 31
признаки, сцепленные с полом........... 16
приспособленность............................... 34
путевой анализ...................................... 27
распределение Пуассона................ 13, 29
решетка Пеннета................................... 36
темп мутирования................................. 30
хромосомная перестройка.................... 28
частота аллеля.......................................... 6
частотно-зависимый отбор 45
эволюционное расстояние................... 47
эффект "бутылочного горлышка"........ 25
эффект "основателя"............................. 25
эффективная численность популяции 24
Гаевский Николай Александрович
Знакомство с эволюционной генетикой
Лицензия ЛР№020372 от 29.01.97
Редактор А.А.Назимова
Корректор Т.Е.Бастрыгина
Подписано в печать 12.09.2002
Усл. печ. л. 3.8
Адрес электронной почты: [email protected]
Издательский центр
Красноярского государственного университета.
660041 Красноярск, пр. Свободный, 79
|
|
|

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Адаптации растений и животных к жизни в горах: Большое значение для жизни организмов в горах имеют степень расчленения, крутизна и экспозиционные различия склонов...

Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...

Индивидуальные очистные сооружения: К классу индивидуальных очистных сооружений относят сооружения, пропускная способность которых...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!