Теоретический анализ роли случайных процессов в популяции, связанных с колебаниями численности составляющих ее особей и частоты встречаемости отдельных генов впервые был сделан Д.Д. Ромашовым (1931). Д.Д. Ромашов и А.С. Серебровский предложили называть эти процессы «автоматическими». Н.П. Дубинин (1931) развил эту идею и ввел термин «генетико-автоматические» процессы. На западе в это же время к аналогичным выводам пришел С. Райт (1931), который ввел ставший в настоящее время распространенным термин «дрейф генов». Позднее понятие о дрейфе генов легло в основу теории нейтральности, сформулированной в 1983 г. М. Кимурой [4].
Дрейф генов относится к особому классу явлений, связанных с “ошибками выборки”. Суть его заключается в том, что генные частоты у ограниченной выборки особей отличаются от таковых у исходной популяции. Например, положив в темный мешок по 50 белых и черных шариков и вытащив наугад только два из них, можно ожидать появления в выборке шариков белого и черного цвета только с вероятностью 0.5. В другой половине случаев в выборке могут оказаться только белые или только черные шарики.
Случайные отклонения исходных генных частот происходят как в большую, так и в меньшую сторону. При этом величина отклонения находится в обратной зависимости от величины выборки. Если выборка состоит из особей, участвующих в воспроизводстве следующего поколения, то чем она меньше, тем больше амплитуда отклонений частот аллелей у потомства по сравнению с родительской популяцией.
Существует понятие "эффективная численность популяции", которая соответствует числу N особей (или числу родителей), дающих начало следующему поколению. Это понятие стало ключевым при рассмотрении эффекта дрейфа генов. В реальных популяциях эффективная численность популяции зависит от структуры скрещивания [4]. Например, если популяция состоит из Nm самцов и Nf самок, участвующих в размножении, эффективный размер популяции равен
 . (24)
. (24)
Если соотношение самцов и самок в популяции 1:1, эффективная численность равна удвоенному значению количества самцов (самок), т.е. вся популяция участвует в размножении. Если соотношение полов неравноценно, то эффективная численность определяется количеством особей менее многочисленного пола. При расчете эффективной численности также следует учитывать возможность передачи разного числа гамет, периодическое изменение числа особей, перекрывание поколений во времени, появление и исчезновение изолированных субпопуляций и другие условия.
Мерой изменчивости генных частот, возникающей в популяции за счет дрейфа генов, служит величина стандартного отклонения:
 , (25)
, (25)
где p и q - частоты аллелей анализируемого локуса; Ne - эффективный размер. В случае, когда количество самцов и самок одинаково (N), выражение (23) имеет вид
 . (26)
. (26)
Максимальная величина s =0.353 при p = q =0.5 и N =1.
Окончательным результатом дрейфа генов является закрепление в популяции одного из аллелей. Вероятность этого события равна вероятности (частоте) распространения аллеля в популяции в начале действия дрейфа генов. Например, если частота аллеля p = 0.5, то равновероятен каждый из исходов, как закрепление аллеля (p = 1) в популяции, так и его потеря (p = 0). Если p = 0.9, то с вероятностью 0.9 этот аллель закрепится в популяции и с вероятностью 0.1 он будет потерян.
Изменение величины стандартного отклонения зависит от числа поколений, на протяжении которых действует дрейф генов:
 , (27)
, (27)
где t - число поколений. При большом t популяция становится гомоаллельной, а стандартное отклонение принимает максимальное значение (s2 = p0q0). Среднее число поколений t, за которое происходит фиксация или потеря аллеля, примерно в четыре раза больше числа родителей в каждом поколении: t = 4 N.
Проявление дрейфа генов можно наблюдать как на одной популяции, так и на их совокупности. При этом одни популяции становятся гомоаллельными по аллелю А, другие по аллелю а, в соответствии с исходным соотношением их частот. Например, при исходных частотах p =0.4, q =0.6 соотношение популяций, в которых закрепился аллель А, и популяций, в которых закрепился аллель а, будет 4:6.
Дрейф генов рассматривают в качестве главного фактора в определении генетического состава популяции, берущей начало от небольшого числа особей (эффект «основателя»и эффект "бутылочного горлышка"), а также при оценке вероятности фиксации селективно нейтральной мутации.
Инбридинг
Одно из главных допущений модели Харди-Вайнберга - случайность скрещивания - в реальных популяциях реализуется очень редко. Ограничение возможностей в выборе партнера в локальной субпопуляции увеличивает родство особей, участвующих в скрещивании. Возникающие близкородственные скрещивания получили название инбридинга. Генетическое значение инбридинга заключается в постоянном увеличении в популяции доли гомозиготных генотипов, в которых встречаются аллели, идентичные не только по функции, но и по происхождению, т.е. берущие начало от общего предка. Редкие рецессивные гены, оказавшись в гомозиготном состоянии, могут проявиться фенотипически. Закрепление в локусе какого-либо одного типа аллеля приводит к появлению чистых линий. Важной особенностью инбридинга является постоянство частот аллелей во всех инбредных поколениях, наблюдаемое на фоне уменьшения частоты гетерозиготных генотипов.
Это можно продемонстрировать на примере популяции с облигатным самооплодотворением (например, у растений самоопылителей). Результаты инбридинга в популяции самоопылителей представлены в табл. 6. В исходном поколении, состоящем из одних гетерозиготных особей, после самооплодотворения появляются гомозиготы (АА, аа). У гомозигот оба аллеля идентичны по происхождению. Доля этих особей в популяции соответствует коэффициенту инбридинга F и составляет после самооплодотворения гетерозигот 0.5. В следующем поколении в результате инбридинга доля гетерозигот вновь уменьшится в два раза и составит ¼, а доля инбредных особей (коэффициент F) возрастет до ¾. Через n поколений доля инбредных особей составит 1-1/2 n , а при бесконечном числе поколений достигнет единицы.
Таблица 6
| Поколение
| Частоты генотипов
| Коэффи-
| Частота
|
|
| АА
| Аа
| аа
| циент. F
| аллеля А
|
| 0 (исходное)
|
|
|
|
| 0.5
|
|
| 1/4
| 1/2
| 1/4
| 0.5
| 0.5
|
|
| 3/8
| 1/4
| 3/8
| 0.75
| 0.5
|
| n
| 0.5· (1-1/2n)
| 1/2n
| 0.5· (1-1/2n)
| (1-1/2n)
| 0.5
|
| ¥
| 1/2
|
| 1/2
|
| 0.5
|
Коэффициент инбридинга F отражает не только потерю гетерозиготности, но и вероятность встречи идентичных по происхождению аллелей в генотипе одной особи. В этом случае величина F зависит от степени генетического родства ее родителей и может быть определена путем анализа ее родословной, которую удобно изображать в виде схемы. Определим коэффициент инбридинга на примере потомства сибсов (родных братьев и сестер). Для этого случая родословная имеет следующий вид:
Родители "А" и "В", по условию, в родстве не состоят. Обозначим аллели гена А у особи "А" - а1а2, а у особи "В" - а3а4. Различные индексы показывают, что аллели не идентичны по происхождению. Вероятность появления у "Е" комбинации а1а1 равна 1/16. Поскольку с такой же вероятностью могут появиться и три другие комбинации (а2а2, а3а3, а4а4), общая вероятность появления у “Е” комбинации идентичных аллелей равна 1/4.
Существует простой способ, называемый "путевым анализом", для определения коэффициента инбридинга по родословной. Он основан на подсчете числа “этапов”, образующих замкнутые циклы, которые включают анализируемую особь и всех предков, общих для обоих родителей. В данном случае это два пути Е-С-А-D-Е и Е-С-В-D-Е по четыре этапа. Поскольку Е появляется дважды, число этапов сокращают на единицу (в нашем примере с четырех до трех). Коэффициент инбридинга определяют как сумму: F = (1/2)3 + (1/2)3 = ¼.
В общем виде, если известно число этапов n, коэффициент инбридинга равен
F = (1/2)n-2. (28)
Установленный коэффициент инбридинга для одной особи справедлив и для одного поколения. Например, популяция, в которой скрещивание происходит между сибсами (F=1/4), в каждом поколении увеличивает долю гомозигот за счет перехода одной четверти генотипов с неидентичным по происхождению набором аллелей в гомозиготное состояние.
Определить долю гомозиготных генотипов Ngg в популяции, находящейся под влиянием инбридинга, можно по формуле
 , (29)
, (29)
где pAa - частота гетерозигот в исходном поколении до начала инбридинга; F - коэффициент инбридинга на одно поколение; t - число поколений инбридинга.
На рис. 4 показано изменение доли гомозиготных генотипов при различных коэффициентах инбридинга в ряду поколений.
Райт включил коэффициент инбридинга в уравнение Харди-Вайнберга для определения соотношения равновесных частот генотипов АА: Аа: аа:
(p2 + F·pq): (2pq - F·´2pq): (q2 + F·pq). (30)
При F= 0.5 частота гетерозигот (2pq) уменьшается на величину pq, т.е. в 2 раза, и соответственно увеличиваются частоты гетерозигот на 0.25 от 2pq. В панмиктической популяции коэффициент F равен нулю и выражение сводится к формуле Харди-Вайнберга.

Рис.4. Доля гомозиготных потомков при систематических скрещиваниях с разной степенью инбридинга
Концепцию идентичности аллелей можно использовать для лучшего понимания смысла выражения Райта, представив частоты генотипов как сумму частот локусов с идентичными и независимыми по происхождению аллелями:
| Генотипы
| Частоты независимых
генотипов
| Частоты идентичных
генотипов
|
| АА
| p2(1-F)
| Fp
|
| Аа
| 2pq(1-F)
| -
|
| аа
| q2(1-F)
| Fq
|
Мутации
Существование мутантных аллелей в популяции связано как с процессом их появления, так и с возможностями их распространения и закрепления.
Сначала рассмотрим судьбу единичной мутации, повторное возникновение которой крайне маловероятно. Примером может служить хромосомная перестройка, частота возникновения которой настолько низка, что ее можно рассматривать как одноразовое событие. Вопрос о судьбе единичной мутации был поставлен и решен Фишером (1930), который предложил в качестве модели популяцию диплоидных размножающихся половым путем организмов. По условиям модели все особи, за исключением одной, гомозиготны по анализируемому локусу АА, а единственная гетерозиготная особь (Аа) одновременно является носителем мутантного аллеля (а), генотип Аа не обладает преимуществом перед исходным генотипом АА, общее количество особей в популяции остается постоянным на протяжении ряда поколений.
Поскольку партнером у гетерозиготы Аа при первом скрещивании всегда будет гомозигота АА, нетрудно показать, каким образом вероятность потери мутантного аллеля (Pп.м.а.) зависит от числа рождаемых потомков n:
| Число потомков n
|
|
|
|
| .....
| n
|
| Вероятность потери гена а
|
| 1/2
| 1/4
| 1/8
| .....
| (1/2)n
|
При неизменной величине популяции среднее число потомков у одной родительской пары равно двум, а возможное число потомков изменяется от нуля до n. Вероятность появления того или иного количества потомков можно описать распределением Пуассона(11), приняв l=2:
 (31)
(31)
Суммируя произведения вероятности потери мутации и вероятности рождения определенного количества потомков, получают вероятность Ploss потери аллеля а за одно поколение:
 (32)
(32)
Расчеты показывают, что эта величина равна e-1 или 0.3679. Вероятность сохранения аллеля а во втором поколении (1 - 0.3679 = 0.6321) выше, чем вероятность его потери. Подобные расчеты для угасающей популяции (среднее число потомков на одну пару - 1, l=1) показывают, что вероятность потери возрастает до 0.606. Наоборот, в случае “процветания”, когда среднее число потомков составляет 3 и 4, вероятность потери снижается до 0.223 и 0.135 соответственно. В любом случае в ряду последующих поколений число редких нейтральных мутаций в популяции будет уменьшаться, пока не исчезнет последний мутантный ген.
В большинстве случаев мутации в одном и том же локусе возникают с определенной частотой или вероятностью. Рассмотрим ситуацию, при которой процесс мутирования является единственным фактором изменения частот аллелей в популяции. Определения частоты (вероятности) появления мутации в популяции понятно из следующего примера. Если за одно поколение мутирует один аллель из 100 тысяч таких аллелей, то частота равна 1/100000=1·10-5. В естественных условиях обычно наблюдаются величины 10-6 - 10-5. Низкий темп мутирования4·10-9 обнаружен у Escherichia soli по гену, определяющему устойчивость к стрептомицину, наиболее высокий - у человека при появлении гена нейрофиброматоза.
При анализе мутационного процесса можно исходить из двух условий: 1 - процесс идет в одном направлении, например ген А трансформируется в ген а; 2 - процесс идет в прямом и обратном направлениях.
Пусть ро частота аллеля А в исходном поколении, u - частота или скорость мутирования аллеля А ® а. Тогда частота аллеля А за одно поколение уменьшится на величину, равную u ´ po. В следующем поколении частота аллеля А составит:
p1 = po - (u · po) = po · (1 - u). (32)
Нетрудно получить выражение для определения частоты аллеля А через t поколений при неизменном темпе мутирования:
pt = po · (1- u)t. (33)
При t стремящемся к бесконечности частота аллеля А (pt)стремится к нулю, или другими словами аллель а полностью замещает аллель А.
Поскольку величина "u" много меньше единицы, выражение (1 - u)t в выражении (33) приближенно равно e -ut. Тогда формула приобретает вид
pt = po · e-ut (34)
Из уравнения 34 легко определить число поколений, необходимое для мутационного сдвига частот аллелей:
 (35)
(35)
Часто определяют число поколений, за которые частота аллеля А уменьшится в 2 раза. Подставив в выражение (35) значение p t = 0.5 po, получим t = ln2/u или t = 0.69/u. При среднем значении u=10-5, t = 69000 поколений.
Для описания процесса мутирования, идущего в прямом и обратном направлении, введем дополнительную переменную v, которая характеризует скорость обратной мутации a ® А. В первом поколении после начала мутаций частота аллеля А составит:
p1 = po - u · po + v · qo или p1 = po - u · po + v · (1- po) (36)
Выражение u · po + v · (1- po) отражает величину изменения частоты аллеля А за одно поколение. В отличие от первого варианта, когда ген мутирует в одном направлении, при одновременном прямом и обратном мутировании величина изменения частоты аллеля А в определенный момент становится равной нулю и наступает динамическое равновесие. Математически это можно представить как: u · pr + v ·(1- pr) = 0, где pr равновесная частота аллеля А. Величина pr = v/(u + v) не зависит от исходной частоты аллеля А и определяется только соотношением скоростей прямого и обратного мутирования.
Таким образом, в большой не подверженной действию отбора популяции мутационный процесс приводит либо к замене одного аллеля другим, либо к равновесию частот. Однако для проявления результатов мутационного процесса из-за малых скоростей требуются большие интервалы времени либо большое число поколений.
Миграция(поток генов)
Межпопуляционный обмен генов возникает при скрещивании особей, принадлежащих разным популяциям. Этому предшествует перемещение особей в пространстве или миграция. В процессе обмена генами можно выделить популяцию, из которой происходят мигранты, и популяцию (аборигены), в которой эти мигранты находят партнера для скрещивания. Если процесс обмена продолжителен, говорят о потоке генов. Эффективность обмена генами зависит от степени или частоты миграции М и от степени генетических различий популяций.
Эволюционная роль миграции не одинакова. При репродуктивном контакте сходных по генетическому составу популяций может происходить как "стирание" существовавших между ними различий, так и сохранение полиморфизма. В большинстве случаев гибридизация между близкими видами не ведет к обмену генами из-за слабой приспособленности потомков. Однако иногда слабо приспособленные потомки достаточно плодовиты, чтобы дать во втором поколении потомство при возвратном скрещивании. Рекомбинантные генотипы от многократных возвратных скрещиваний будут все более и более приближаться к родительскому типу и могут влиться в генофонд родительской линии. В результате этого некоторое количество отдельных генов или генных блоков перейдет от одного вида к другому.
Рассмотрим влияние миграции на генетический состав популяции аборигенов. Предположим, что миграция происходит в одном направлении из большой материковой популяции в маленькую островную; все допущения модели Харди-Вайнберга остаются в силе; поток генов происходит с постоянной скоростью М, рассчитанной как доля мигрантов в популяции аборигенов на одно поколение; частоты аллеля а в материковой и островной популяциях не равны и составляют Q и q соответственно; за время миграции частота аллеля а в материковой популяции остается постоянной, так как число мигрантов мало по сравнению с численностью материковой популяции.
Рассмотрим численный пример. Пусть частота аллеля а в материковой популяции равна Q = 0.7, в основной - до начала миграции qо= 0.4; за одно поколение в островную популяцию вливается 20%. После слияния островная популяция будет представлена мигрантами (M = 0.2) и аборигенами (1- М = 0.8). Частота аллеля а при этом составит q1 = 0.2·0.7+0.8·0.4 = 0.46. В общем виде значение частоты аллеля а в первом поколении можно определить следующим образом:
q1 = (1 - M) ·qo + M·Q = qo - M·(qo - Q). (37)
Выражение [- M·(qo - Q)] отражает изменение частоты аллеля а за одно поколение. Оно обращается в ноль, когда прекращается процесс миграции (М = 0), либо когда частоты аллеля а в обеих популяциях становятся одинаковыми (q = Q).
Полезно проследить за изменением различий в частотах аллеля а между островной и материковой популяциями в нескольких поколениях при постоянной скорости миграции. В первом поколении различия составят:
q1 - Q = qo - M·(qo - Q) - Q = qo - Mqo + MQ - Q = (1 - M)·qo - (1 - M) ·Q, (38)
q1 - Q = (1- M) · (qo - Q). (39)
Для второго поколения имеем q2 = q1 - M·(q1 - Q). Подставив вместо q1 выражение (37), получаем:
q2 =(1 - M)2·qo + 2M·Q - M2·Q.
Различия частот во втором поколении составят:
q2 - Q = (1 - М)2 ·(qo - Q).
Очевидно, что через t поколений это выражение примет вид:
qt - Q = (1 - М)t ·(q0 - Q). (40)
Эта формула дает возможность рассчитать частоту аллеля в локальной популяции по прошествии t поколений с начала миграции, если известны исходные частоты аллеля qo и Q и скорость миграции М:
qt = (1 - М)t · (q0 - Q) + Q (41)
либо скорость миграции (t) и долю генов предков (1-М)t в популяции аборигенов по прошествии t поколений:
(1 - М)t = (qt - Q)/(q0 - Q) (42)
Воспользуемся примером из [1]. В США потомство от смешанных браков между белыми и неграми принято относить к негритянскому населению. Поэтому смешанные браки можно рассматривать как поток генов из белой популяции в темнокожую. Предки современных негров в США были переселены из Африки около 300 лет назад, что соответствует 10 поколениям (t=10). Исходные значения частот аллелей гена q0, кодирующего белок крови, можно получить на основе анализов крови у современных чернокожих, живущих в Африке. Для оценки современных частот pt подойдут чернокожие штатов Джорджия и Калифорния, величину Р дадут белые США.
Проанализируем возможные величины потока генов М от белого населения к темнокожему и долю генов негритянских предков у современных негров на основе частот аллелей, определяющих свойства крови. Сравним популяции чернокожих двух указанных штатов, а также однозначность оценок при использовании в качестве маркеров различных аллелей. Оценим разброс данных и определим достоверность различий средних по данному параметру у двух популяций темнокожих людей. Исходные данные представлены в табл. 7.
Таблица 7
| Аллель
| Темнокожие Африки
(po,q0)
| Темноко-жие Джорджии
(pt,qt)
| Темноко-жие Калифорнии
(pt,qt)
| Белые США
(Р,Q)
| М,
для Джорд-жии
| М,
для Калифор-нии
|
| Ro
| 0.630
| 0.533
| 0.486
| 0.022
| 0.017
| 0.027
|
| R1
| 0.066
| 0.109
| 0.161
| 0.429
| 0.013
| 0.030
|
| R2
| 0.061
| 0.069
| 0.071
| 0.137
| 0.011
| 0.014
|
| r
| 0.238
| 0.250
| 0.253
| 0.374
| 0.009
| 0.012
|
| A
| 0.156
| 0.166
| 0.175
| 0.241
| 0.012
| 0.025
|
| B
| 0.136
| 0.113
| 0.125
| 0.038
| 0.026
| 0.012
|
| M
| 0.474
| 0.484
| 0.486
| 0.507
| 0.035
| 0.044
|
| S
| 0.172
| 0.175
| 0.181
| 0.279
| 0.003
| 0.009
|
| Среднее и его ошибка:
| 0.016
±0.003
| 0.021
±0.004
|
Расчеты показывают, что в штате Джорджия поток генов от белого населения к темнокожему за одно поколение составляет 1.6%, в штате Калифорния - 2.1%. Различия заметны, но недостоверны (tst = 0.96). Доля генов предков у негритянского населения США после 10 поколений в штатах Джорджия и Калифорния соответственно равны 0.851 и 0.809.
Естественный отбор
Между особями, составляющими вид, существует множество различий. Особи в популяции, которые обладают признаками лучшего выживания, оставляют больше потомства и считаются более приспособленными. Главная особенность приспособительного признака состоит в том, что он тем или иным способом повышает шансы особи участвовать в создании популяции следующего поколения. Результатом естественного отбора может быть создание, сохранение или потеря изменчивости в популяции. Отбор генов или генных комплексов осуществляется на уровне их фенотипических проявлений. Аллели, способствующие увеличению приспособленности фенотипа, также подвержены отбору на их сохранение. Поэтому силу отбора можно выразить в терминах превосходства аллелей и изменениях частот генов, хотя в действительности конкурируют не гены, а особи и их группы.
При описании отбора используют такие показатели как адаптационная ценность генотипаили приспособленностьW и коэффициент отбораs. Теоретически адаптационная ценность должна выражать как непосредственную приспособленность генотипа, которую можно оценить через число реальных потомков, так и долгосрочную - вклад этих генотипов в последующие генофонды. В реальной ситуации долгосрочную приспособленность можно оценить на основе формализованной математической модели.
Адаптационная ценность (приспособленность) каждого генотипа зависит от взаимодействия всех генов, имеющихся у данной особи. Если селективная ценность всех генотипов, содержащих аллель а1, больше селективной ценности всех генотипов с аллелем а2, отбор будет направлен на сохранение аллеля а1, частота которого из поколения в поколение будет расти.
Модель отбора генотипов на уровне одного аутосомного локуса. Местом действия отбора выберем панмиктическую популяцию. Предпосылкой отбора выступают различия в адаптационной ценности генотипов (фенотипов), которые возникают в промежуток времени с момента первого деления зиготы и до образования зиготы следующего поколения. Если некий локус расщепляется по аллелям А и а, популяцию можно разделить по генотипам на три группы. В каждой группе генотип имеет свою адаптационную ценность АА - Wo; Аа - W1, аа - W2. Положим, что при образовании поколения зигот, принятого за точку отсчета, частоты генотипов у самцов и самок были одинаковы и составляли для АА – x, для Аа – y и для аа – z. Сумма частот равна единице x+y+z=1.
Коэффициент участия каждого генотипа в создании следующего поколения зигот определяется произведением частоты генотипа на показатель его адаптационной ценности (или приспособленности):
| Генотипы
| АА
| Аа
| аа
| Сумма
|
| Коэффициент
участия
| x·Wo
| y·W1
| z·W2
| <1
|
Поскольку показатели адаптационной ценности принято нормировать по максимальному значению одного из коэффициентов Wo , W1 , W2, получаемая сумма меньше единицы.
Частоты генотипов, участвующих в образовании зиготы следующего поколения определяются следующими выражениями:
 , (43)
, (43)
где  представляет собой среднюю приспособленность популяции по анализируемому локусу.
представляет собой среднюю приспособленность популяции по анализируемому локусу.
Частоты аллелей в этой выборке генотипов имеют значения
 (44)
(44)
При условии, что характеристики субпопуляций самцов и самок совпадают, для определения частот генотипов образовавшихся зигот следующего поколения можно воспользоваться выражением Харди-Вайнберга:
 (45)
(45)
Предложенный принцип позволяет продолжить определение частот аллелей и генотипов в последующих поколениях зигот и проследить их динамику в условиях действия отбора.
К уравнениям (45) можно прийти и другим путем, соотнося частоты генотипов потомков с частотами различных вариантов скрещиваний. При условии, что характеристики субпопуляций самцов и самок совпадают, следует ожидать, что различные варианты скрещивания (в соответствии с решеткой Пеннета) будут возникать пропорционально частоте и приспособленности генотипов.
В рамках обсуждения результатов отбора необходимо рассмотреть изменение средней приспособленности популяции за один этап отбора. Дисперсия приспособленности в начальный момент времени равна
 . (46)
. (46)
Раскрыв скобки с учетом того, что p2+2pq+q2=1, получим
 . (47)
. (47)
После отбора средняя приспособленность популяции будет
 . (48)
. (48)
Объединяя формулы (47) и (48), получаем
 . (49)
. (49)
Изменение средней приспособленности за поколение составляет
 (50)
(50)
На основании полученного результата Р.Фишер в 1930 г. сформулировал “фундаментальную теорему естественного отбора”: Изменение средней приспособленности прямо пропорционально ее дисперсии в начальный момент времени и обратно пропорционально ее величине.
Результат действия отбора за одно поколение оценивают по изменению частот аллелей. Например, изменение частоты аллеля а можно представить как
 . (51)
. (51)
Для случая, когда в исходном поколении частоты генотипов соответствовали уравнению Харди-Вайнберга, необходимо в выражении (51) произвести следующие подстановки: x = p2, y = 2pq, z = q2. После ряда несложных математических преобразований выражение (51) приобретает вид
 . (52)
. (52)
Знаменатель, как уже отмечалось, представляет собой среднюю приспособленность популяции при действии отбора и обозначается Ws.
Поскольку относительные величины коэффициентов приспособленности, не выходят за пределы интервала 0£W£1, в выражении (52) можно заменить коэффициенты адаптационной ценности на соответствующие значения коэффициентов отбора si, принимая, что Wi+si=1 (Wo = 1- so, W1 = 1 - s1,W2 = 1- s2). Тогда
 . (53)
. (53)
Средняя приспособленность популяции при этом имеет вид:
Ws = 1 - (p2so + 2pqs1 + q2s2). (54)
Следует заметить, что оценка изменения частоты аллеля справедлива только по отношению к исходной популяции и первому поколению потомков, возникшему в условиях действия отбора. Оно не отражает изменения частоты аллеля в целом, поскольку не определены такие параметры, как возрастной состав популяции, длительность репродуктивного периода, доля потомков и т.п.
В рамках полученной модели (52) изменение частоты аллеля а в ходе отбора прямо пропорционально доле гетерозигот pq и обратно пропорционально средней приспособленности популяции Ws. Часть выражения (52) - [q(W2 -W1) + p(W1 - Wo)] наиболее важна при анализе последствий отбора. Она состоит из двух слагаемых, в которых сопоставляются приспособленности генотипов при последовательном вытеснении одного аллеля другим (аа -> Аа -> АА), что в конечном итоге определяет направление (знак) действия отбора.
Модель отбора позволяет определить условия, при которых не происходит изменение частот аллелей [5]. Согласно (52) при этом q1- q0=0. Кроме “краевых условий”, соответствующих завершению отбора после полного вытеснения одного аллеля другим (p=1, q=0 или p=0, q=1), существует третье равновесное состояние популяции, при котором q(W2‑W1) + p(W1‑Wo) = 0. Решая это уравнение относительно p, получим
 . (55)
. (55)
Условие, когда 0 < p < 1 может выполняться в двух случаях:
1) W2 -W1>0, W2‑2W1+W0>0, W2‑2W1+W0>W2 -W1;
2) W2 -W1<0, W2‑2W1+W0<0, W2‑2W1+W0<W2 -W1.
В первом - решение неравенств дает W2>W1, W0>W1, т.е. приспособленность гетерозигот меньше приспособленности гомозигот (отбор против гетерозигот), во втором - W2<W1, W0<W1 - приспособленность гетерозигот выше, чем приспособленность гомозигот (отбор в пользу гетерозигот). Ниже мы определим в каком из этих двух случаев равновесие устойчивое, а в каком – нет.
При анализе различных ситуаций, связанных с отбором генотипов, необходимо учитывать возможность нулевого, частичного и полного доминирования одного аллеля над другим и степень отбора (от частичного до полного, когда генотипы летальны или стерильны). Можно рассмотреть пять основных вариантов отбора (табл. 8), определив для каждого коэффициенты относительной приспособленности (Wi = 1- si) и условия доминирования.
Таблица 8
| №
| Условия отбора
| Относительная приспособленность генотипа Wi
| Аналитическое
выражение для оценки
|
|
|
| AA(W0)
| Aa(W1)
| aa(W2)
| Dq= q1-q0
|
|
|
|
|
|
|
|
|
| Полное доминирование, отбор против аа
|
1
|
1
|
1-s
| 
|
|
| Полное доминирование, отбор против АА и Аа
|
1-s
|
1-s
|
1
| 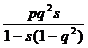
|
|
| Отсутствие доминирования
| 1-s0*
| 1-s1**
| 1
| 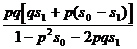
|
|
| Сверхдоминирование отбор против АА и аа
| 1-s0
| 1
| 1-s2
| 
|
|
| Отбор против гетерозигот
| 1
| 1-s
| 1
| 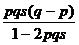
|
| | | | | | | |
*Присвоение коэффициенту отбора соответствующего индекса оправдано в случае, когда приспособленность по крайней мере двух генотипов различна.
** В некоторых случаях можно обойтись одним коэффициентом отбора, если известно соотношение между s0 и s1, например s0=2s1.
Интенсивность отбора и средняя приспособленность популяции зависят не только от коэффициентов отбора, но и от частот отбираемых аллелей. Показательными в этом отношении являются графики зависимости Dq = f(q) и Ws=f(q), с помощью которых можно определить экстремальные и равновесные условия (рис. 5 - 8).
Как было показано выше, при сверхдоминировании и отборе против гетерозигот возможны соответствующие "нетривиальные решения":
 и
и  .
.
Видно, что при сверхдоминировании равновесие наступает при условии
 (56)
(56)
или  , (57)
, (57)
а при отборе против гетерозигот, когда q = p = 0.5.
Знак величины Dq, полученный при значениях q вблизи точки равновесия (аналитически или на основе графика), позволяет определить, является ли равновесие устойчивым или неустойчивым.
Например, при сверхдоминировании (рис.5) равновесие устойчивое, так как отбор увеличивает частоту аллеля а при значениях частот меньше равновесных и, наоборот, уменьшает - при значениях больше равновесных. Как видно из рис. 6, через определенное число поколений в популяции устанавливаются постоянные значения частот генотипов и аллелей. Результат действия отбора можно обнаружить только при условии, если популяцию вывести из состояния равновесия. Отбор в пользу гетерозигот восстановит равновесные частоты. Эта равновесная популяция в условиях отбора отличается от равновесной популяции Харди-Вайнберга, в которой случайное отклонение от равновесия приводит к дрейфу генов.
В варианте отбора против гетерозигот равновесие неустойчивое (рис. 7). Если исходные частоты аллелей не соответствуют равновесным (p=q=0.5), отбор завершается либо полной потерей аллеля а, либо его сохранением в гомозиготном варианте (аа) (рис.8).
Интересно наряду с изменением частот аллелей проследить за поведением параметра Ws, характеризующего среднюю относительную приспособленность популяции. Приспособленность стремится к единице по мере удаления отбором неблагоприятного аллеля и несущих этот аллель генотипов в первых трех рассмотренных в таблице вариантах. При сверхдоминировании приспособленность максимальна, но не равна единице в точке устойчивого равновесия (рис.5). В случае отбора против гетерозигот (рис.8) Ws - минимальна в точке неустойчивого равновесия.

Рис.5. Зависимость скорости отбора D q и средней приспособленности Ws от частоты аллеля а. В уравнениях (53) подставлены следующие коэффициенты: s0 = 0.8, s1 = 0, s2 = 0.7 (отбор в пользу гетерозигот)

Рис.6. Изменения частот г








