Каждая мембрана клетки замкнута, т.е. имеет внутреннюю и внешнюю поверхности, различающиеся по липидному и белковому составам - эту особенность мембран называют трансмембранной (поперечной) асимметрией.
Липидная асимметрия возникает прежде всего потому, что липиды с более объёмными полярными "головками" стремятся находиться в наружном монослое, так как там площадь поверхности, приходящаяся на полярную "головку", больше. Фосфатадилхолины и сфингомиелины локализованы преимущественно в наружном монослое, а фосфатидилэтаноламины и фосфатидилсерины в основном во внутреннем.
Липиды в некоторых биологических мембранах с довольно большой частотой мигрируют с одной стороны мембраны на другую, т.е. совершают "флип-флоп" (от англ, flip-flop) перескоки (рис. 5-7). Перемещение липидных молекул затрудняют полярные "головки", поэтому липиды, находящиеся на внутренней стороне мембраны, имеют относительно высокую скорость трансмембранной миграции по сравнению с липидами наружной стороны мембраны, мигрирующих медленнее или вообще не совершающими "флип-флоп" перескоки.
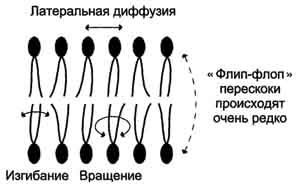
Рис. 5-7. Типы движений липидных молекул в бислое мембран.
Жидкостностъ мембран
Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной диффузии. Скорость перемещения молекул зависит от микровязкости мембран, которая, в свою очередь, определяется относительным содержанием насыщенных и ненасыщенных жирных кислот в составе липидов. Микровязкость меньше, если в составе липидов преобладают ненасыщенные жирные кислоты, и больше при высоком содержании насыщенных жирных кислот.
Ацильные (алифатические) остатки ненасыщенных жирных кислот имеют так называемые "изломы" (см. раздел 8). Эти "изломы" препятствуют слишком плотной упаковке молекул в мембране и делают её более рыхлой, а следовательно и более "текучей". На текучесть мембран также влияют размеры углеводородных "хвостов" липидов, с увеличением длины которых мембрана становится более "текучей".
Функции мембранных липидов
Фосфо- и гликолипиды мембран, помимо участия в формировании липидного бислоя, выполняют ряд других важных функций.
Липиды формируют среду для функционирования Мембранных белков, принимающих в ней нативную конформацию. Выделенные из мембран ферменты, лишённые липидного окружения, как правило, не проявляют каталитической активности.
Некоторые мембранные липиды - предшественники вторичных посредников при передаче гормонального сигнала. Так, фосфати-дилинозитол-4,5-бисфосфат (ФИФ2) под действием фермента фосфолипазы С гидролизуется до диацилглицерола (ДАТ), активатора протеинкиназы С и инозитол-1,4,5-трифосфата (ИФ3) - регулятора кальциевого обмена в клетке (рис. 5-8). ДАГ, ИФ3, протеинкиназа С и Са2+ - участники инозитолфосфатной системы передачи сигнала.
Кроме того, некоторые липиды выполняют "якорную" функцию, например к фосфатидил-инозитолам через олигосахарид могут присоединяться специфические белки наружной поверхности клетки. Фосфатидилинозитол с присоединённым к нему олигосахаридом (гликаном) называют фосфатидилинозитолгликаном. Связь белков с этой молекулой (гликаном) осуществляется через фосфоэтаноламин.

Рис. 5-8. Гидролиз фосфатидилинозитол-4,5-бисфосфата.
Пример такого "заякоренного" белка - ацетилхолинэстераза, катализирующая гидролиз ацетилхолина в синаптической щели. Этот фермент фиксируется на поСтеинаптической мембране, ковалентно присоединяясь к фосфатидилино-зитолгликану. Под действием фосфолипазы С может происходить отделение белков от внешней поверхности клетки.
Липиды могут быть аллостерическими активаторами мембранных ферментов. Например, р-гидроксибутиратдегидрогеназа, участвующая в окислении кетоновых тел (см. раздел 8), локализована на внутренней мембране митохондрий. Каталитическая активность фермента проявляется только в присутствии фосфатидилхолина.
Фермент протеинкиназа С катализирует реакции фосфорилирования белков по аминокислотным остаткам серина и треонина. В неактивной форме протеинкиназа С находится в цитозоле. Однако после стимуляции клетки (повышение в клетке концентрации кальция) фермент быстро активируется ионами кальция и оказывается связанным с мембраной. Функционально активная протеинкиназа С - комплекс, содержащий мономер фермента, молекулу диацилглицерола, один или более ионов Са2+ и четыре молекулы фосфатидилсерина.
Креатинкиназа, фермент катализирующий образование макроэргического соединения креа-тинфосфата (см. раздел 9). Для проявления его активности требуется специфическое взаимодействие с кардиолипином внутренней мембраны митохондрий.
Липидный состав мембран - фосфолипиды, гликолипиды, холестерин. Белки мембран - интегральные, поверхностные, «заякоренные». Роль отдельных компонентов мембран в формировании структуры и выполнении функций.
Липидам принадлежит главная роль в образовании мембран как клеточных структур: пластинчатая, «мембранная» форма и основные физико-химические свойства мембран определяются именно липидами. Основная часть липидов (до 90 %) в мембранах представлена фосфолипидами, гликолипидами и холестерином.
Фосфолипиды. В мембранах имеются фосфолипиды двух типов — глицерофосфолипиды и сфингофосфолипиды.
Глицерофосфолипиды. Эти липиды являются производными фосфатидной кислоты (диацилглицеринфосфата).

 В состав фосфолипидов входят жирные кислоты, как насыщенные, так и ненасыщенные. Наиболее распространенные глицерофосфолипиды — это фосфатидилхолины. Их отличительной чертой является наличие в молекуле остатка холина, связанного с фосфорной кислотой. Фосфатидилхолинами называют группу соединений, отличающихся друг от друга природой жирно-кислотных остатков (радикалов R). По такому же типу построены и другие глицерофосфолипиды — фосфатидилэтаноламины и фосфатидилсерины, содержащие соответственно этаноламин и серии.
В состав фосфолипидов входят жирные кислоты, как насыщенные, так и ненасыщенные. Наиболее распространенные глицерофосфолипиды — это фосфатидилхолины. Их отличительной чертой является наличие в молекуле остатка холина, связанного с фосфорной кислотой. Фосфатидилхолинами называют группу соединений, отличающихся друг от друга природой жирно-кислотных остатков (радикалов R). По такому же типу построены и другие глицерофосфолипиды — фосфатидилэтаноламины и фосфатидилсерины, содержащие соответственно этаноламин и серии.
Специальные функции в мембранах выполняет фосфатидилинозитол, в состав которого входит шестиатомный циклический спирт инозитол. Этот фосфолипид участвует в одном из механизмов передачи внешних регуляторных сигналов через клеточную мембрану в клетку.
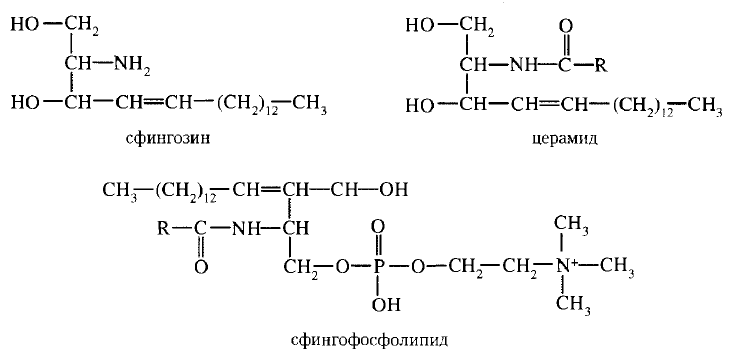 Сфингофосфолипиды (сфингомиелины). В эту группу входят липиды, содержащие аминоспирт сфингозин. Сфингофосфолипиды являются производными церамидов (N-ацилсфингозинов); в остальном сфингофосфолипиды построены сходно с глицерофосфолипидами.
Сфингофосфолипиды (сфингомиелины). В эту группу входят липиды, содержащие аминоспирт сфингозин. Сфингофосфолипиды являются производными церамидов (N-ацилсфингозинов); в остальном сфингофосфолипиды построены сходно с глицерофосфолипидами.
Церамиды — это группа соединений, различающихся по остатку жирной кислоты. В сфингофосфолипидах водород гидроксильной группы у первого углеродного атома церамида замещен на фосфохолин, фосфоэтаноламин или фосфосерин.
Гликолипиды представляют собой углеводсодержащие соединения, в которых углеводная часть ковалентно связана с липидной.
В мембранах содержатся главным образом углеводные производные церамида
(N-ацилсфингозина). Общее название таких гликолипидов — гликозилцерамиды, гликосфинголипиды (их называют также цереброзидами).
Напомним, что приставки глюко- или глюкозил- означают глюкозный остаток, а глико- и гликозил- — любой углеводный остаток. Углеводная часть в гликозилцерамидах может быть представлена моносахаридами или олигосахаридами, например остатком лактозы (дисахарид) в лактозилцерамиде.
В меньших количествах в гликолипидах обнаруживаются углеводы более сложного строения, образующие разветвленные цепи. Они содержат до 20-30 моносахаридов. Концевые остатки углеводных цепей часто представлены N-ацетилней-раминовой кислотой (девятиуглеродный моносахарид); гликолипиды, содержащие N-ацетилнейраминовую кислоту, называют ганглиозидами.
 Холестерин — это представитель группы липидов, называемых стероидами. Характерные черты структуры холестерина — наличие тетрациклической группировки и углеводородной разветвленной цепи с восемью углеродными атомами; в третьем положении полициклической части имеется спиртовая группа.
Холестерин — это представитель группы липидов, называемых стероидами. Характерные черты структуры холестерина — наличие тетрациклической группировки и углеводородной разветвленной цепи с восемью углеродными атомами; в третьем положении полициклической части имеется спиртовая группа.
Белки мембран. Мембранные белки, контактирующие с гидрофобной частью липидного бислоя, должны быть амфифильными. Те участки белка, которые взаимодействуют с углеводородными цепями жирных кислот, содержат преимущественно неполярные аминокислоты. Участки белка, находящиеся в области полярных "головок", обогащены гидрофильными аминокислотными остатками.
Белки мембран различаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его - интегральные белки, либо разными способами прикрепляться к мембране - поверхностные белки.
Поверхностные белки. П рикрепляются к мембране, взаимодействуя с интегральными белками или поверхностными участками липидного слоя.
Белки, образующие комплексы с интегральными белками мембраны. Ряд пищеварительных ферментов, участвующих в гидролизе крахмала и белков, прикрепляется к интегральным белкам мембран микроворсинок кишечника.
Примерами таких комплексов могут быть сахаразаизомальтаза и мальтазагликоамилаза. Возможно, связь этих пищеварительных ферментов с мембраной позволяет с высокой скоростью гидролизовать субстраты и усваивать продукты гидролиза клеткой.
Белки, связанные с полярными «головками» липидов мембран. Полярные или заряженные домены белковой молекулы могут взаимодействовать с полярными "головками" липидов, образуя ионные и водородные связи. Кроме того, множество растворимых в цитозоле белков при определённых условиях могут связываться с поверхностью мембраны на непродолжительное время. Иногда связывание белка -- необходимое условие проявления ферментативной активности. К таким белкам, например, относят протеинкиназу С, факторы свёртывания крови.
Закрепление с помощью мембранного "якоря"
"Якорем" может быть неполярный домен белка, построенный из аминокислот с гидрофобными радикалами. Примером такого белка может служить цитохром b5 мембраны ЭР. Этот белок участвует в окислительно-восстановительных реакциях, как переносчик электронов.
Роль мембранного "якоря" может выполнять также ковалентно связанный с белком остаток жирной кислоты (миристиновой - С14 или пальмитиновой - С16). Белки, связанные с жирными кислотами, локализованы в основном на внутренней поверхности плазматической мембраны. Миристиновая кислота присоединяется к N-концевому глицину с образованием амидной связи. Пальмитиновая кислота образует тиоэфирную связь с цистеином или сложноэфирную с остатками серина и треонина.
Небольшая группа белков может взаимодействовать с наружной поверхностью клетки с помощью ковалентно присоединённого к С-концу белка фосфатидилинозитолгликана. Этот "якорь" - часто единственное связующее звено между белком и мембраной, поэтому при действии фосфолипазы С этот белок отделяется от мембраны.








