Характерен для нервной ткани. Встречается между двумя нейронами или между нейроном и рецептором/эффектором (к примеру, нервно-мышечное окончание). Синапсы - участки контактов двух клеток, специализированных для односторонней передачи возбуждения/торможения. Синапсы - это терминальные участки дендритов и аксонов, образующие связь как с телом другой нервной клетки, так и с её отростками. Мембраны этих клеток разделены синаптической щелью 20-30 нм. Мембрана в области синаптического контакта одной клетки - пресинаптическая, а другой, воспринимающей импульс, - постсинаптической. Около пресинаптической мембраны много синаптических пузырьков, заполненных медиаторами. В момент прохождения нервного импульса синаптические пузырьки выбрасывают своё содержимое в синаптическую щель.
Плазмодесмы
Характерны для растений.
Тонкие трубчатые цитоплазматические каналы диаметром 20-40 нм, соединяющие две соседние клетки. Ограничивающая эти каналы мембрана непосредственно переходит в ПМ соседствующих клеток. Плазмодесмы проходят сквозь клеточную стенку двух соседних клеток и соединяют их гиалоплазму. Это напоминает синцитий: объединение многих клеточных территорий с помощью цитоплазматических мостиков. Плазмодесмы образуются во время деления клетки, когда строится первичная клеточная стенка.
Функция плазмодесм: межклеточная циркуляция растворов, липидных капель, вирусов. Свободный транспорт через плазмодесмы ограничивается частицами с массой не более 800 дальтон.
КЛЕТОЧНАЯ СТЕНКА (ОБОЛОЧКА) РАСТЕНИЙ
Если выделить из организма животного клетку и поместить её в воду, она набухнет, лопнет, лизируется в рез-те проникновения в цитоплазму нек. солей. В организме этого не происходит, т.к. животные клетки окружены жидкостями внутренней среды. Свободноживущие в пресной воде простейшие, у которых нет КС, не лизируются за счёт постоянной работы сократительных вакуолей. Если поместить в воду клетки бактерий и растений, они не будут лизироваться до тех пор, пока цела их КС, которую можно растворить различными ферментами.
КС растений формируется при участии ПМ и является экстраклеточным (внеклеточным) многослойным образованием, как бы служащим наружным скелетом растительной клетки. КС растений состоит из двух компонентов: из аморфного пластичного гелеобразного матрикса (основы), содержащего много воды, и опорной фибриллярной системы. К тому же, в состав КС могут входить доп. в-ва и соли для придания ей жёсткости, несмачиваемости и т.д. К примеру, лигнин приводит к одревеснению КС, повышению их прочности.
В состав матрикса КС растений входят полисахариды, гемицеллюлозы и пектиновые в-ва. Гемицеллюлозы - ветвящиеся полимерные цепи гексоз (глюкоза, манноза, галактоза и др.), пентоз (ксилоза, арабиноза) и уроновых кислот (глюкуроновая и галактуроновая). Пектиновые вещества - разветвлённые полимеры, несущие отрицательные заряды из-за множества остатков галактуроновой кислоты. Гемицеллюлозы и пектины синтезируются в вакуолях АГ.
Целлюлоза - линейный неветвящийся полимер β-глюкозы, образующий в КС растений фибриллы (целлюлоза --> субмикроскопические микрофибриллы толщиной 25 нм --> фибриллы). Фибриллы целлюлозы синтезируются специальными ферментами, встроенными в плазмалемму. Целлюлоза (клетчатка) - самый распространённый по массе биополимер на Земле. В ней аккумулировано около 50% всего углерода живого вещества биосферы. Только некоторые микроорганизмы способны расщеплять целлюлозы. В клетках животных и растений таких ферментов нет.
На поверхностях КС могут накапливаться различные адкрустирующие в-ва (кутин, суберин), приводящие к опробковению клеток. В клетках эпидермиса на поверхности КС откладывается воск, образующий водонепроницаемый слой --> препятствует потере клеткой воды.
Образование фрагмопласта (клеточной пластинки): при делении клеток растений после расхождения хромосом в экваториальной плоскости появляется скопление мелких мембранных пузырьков, которые начинают сливаться друг с другом по направлению от центра клетки к периферии.
ПРОЦЕСС ОБРАЗОВАНИЯ КЛЕТОЧНОЙ СТЕНКИ РАСТЕНИЙ
КС растений формируется при участии ПМ и является экстраклеточным (внеклеточным) многослойным образованием, защищающим поверхность клетки, служащим как бы наружным скелетом растительной клетки. При росте КС выделение в-в, из к-х она строится, происходит по всей поверхности клетки. Аморфные в-ва матрикса (гемицеллюлозы и пектины) синтезируются в вакуолях АГ и выделяются через плазмалемму путём экзоцитоза. Фибриллы целлюлозы синтезируются спец. ферментами, встроенными в плазмалемму.
???? http://www.mylect.ru/biology/botanika/647-stroenie-ra.html?start=8
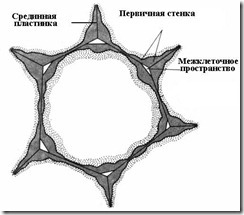
Образуется фрагмопласт (клеточная пластинка), содержащий пектиновые в-ва, нет целлюлозы. Во взрослой клетке клеточная пластинка претерпевает изменения, становится срединной (межклеточной) пластинкой.
Сразу после образования клеточной пластинки протопласты дочерних клеток начинают откладывать свою собственную клеточную стенку. Она откладывается изнутри как на поверхности клеточной пластинки, так и на поверхности других клеточных стенок, принадлежавших ранее материнской клетке. Клетка растёт засчёт осмоса, роста центр. вакуоли. Тургорное давление начинает растягивать стенку, но она не рвется благодаря тому, что в нее постоянно откладываются новые порции микрофибрилл и веществ матрикса. Стенки делящихся и растущих клеток называют первичными - содержат целлюлозу, но не содержат лигнин. Для многих клеток отложение клеточной стенки прекращается одновременно с прекращением роста клетки. Такие клетки окружены тонкой первичной стенкой до конца жизни. У других клеток отложение стенки продолжается и по достижении клеткой окончательного размера. При этом толщина стенки увеличивается, а объем, занимаемый полостью клетки, сокращается. Такой процесс носит название вторичного утолщения стенки, а саму стенку называют вторичной . Во вторичной стенке преобладают фибриллы целлюлозы.
Срединная пластинка склеивает соседние клетки. Если ее растворить, стенки клеток теряют связь друг с другом и разъединяются. Этот процесс называется мацерация. Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой (перезрелые плоды груши, дыни, персика, банана). Часто наблюдается частичная мацерация, при которой срединная пластинка растворяется не по всей поверхности, а лишь в углах клеток. Вследствие тургорного давления соседние клетки в этих местах округляются, в результате чего образуются межклетники (рис. 2.16 ). Межклетники образуют единую разветвленную сеть, которая заполняется парами воды и газами. Таким образом, межклетники улучшают газообмен клеток.
ВСПОМНИТЬ ПРО ПОРЫ ИЗ ВЫСШРАСТА!!!
При делении клеток растений после расхождения хромосом в экваториальной плоскости появляется скопление мелких мембранных пузырьков, которые начинают сливаться друг с другом по направлению от центра клетки к периферии - образование клеточной пластинки (фрагмопласта). В центральной части фрагмопласта располагается аморфное в-во матрикса, которое наполняло сливающиеся пузырьки. Эти первичные вакуоли происходят от мембран АГ. В состав первичной КС входит также небольшое кол-во (около 10%) белка, богатого гидроксипролином. По периферии фрагмопласта при наблюдении её в поляризованном свете обнаруживается двойное лучепреломление, вызванное тем, что в этом месте располагаются ориентированные фибриллы целлюлозы. Таким образом, растущая КС состоит из трёх слоёв: центральная - срединная пластинка только из аморфного матрикса, два периферических - первичная оболочка, содержащая гемицеллюлозу и целлюлозные фибриллы. Срединная пластинка - продукт активности исходной клетки. Первичная оболочка - образуется за счёт выделения гемицеллюлозы и фибрилл целюлозы двумя новыми клеточными телами. Далее происходит выделение веществ матрикса по тому же механизму. Затем вне клетки на её ПМ идёт синтез и полимеризация целлюлозных фибрилл. Так постепенно образуется вторичная КС, которая соединена с первичной несколькими промежуточными слоями. Вторичная КС придаёт клетке её окончательную форму. После разделения клетки на две происходит рост новых клеток, увеличение их объёма, изменение формы.
В первичной КС фибриллы могли скользить относительно друг друга. В дальнейшем при замещении в матриксе гемицеллюлозы на лигнин подвижность фибрилл резко снижается, оболочка становится плотной, поисходит одревеснение. Часто под вторичной КС обнаруживают третичную - засохший остаток дегенерировавшего слоя собственного цитоплазмы.
ДОПИСАТЬ
КЛЕТОЧНЫЕ ОБОЛОЧКИ БАКТЕРИЙ
КС бактерий состоит из муреина - пептидогликана. Жёсткий каркас, окружающий бактериальную клетку, называется муреиновым мешком. Основа его структуры - сеть параллельных полисахаридных цепей из чередующихся дисахаридов (ацетилглюкозамин, соединённый с ацетилмурамовой кислотой). Основа пептидной части муреина - тетрапептиды из а/к.
При окраске по Граму (окраска кристаллическим фиолетовым, обработка иодом, отмывка спиртом) бактерии по-разному воспринимают краситель: Гр+ остаются окрашенными после обработки спиртом, Гр- обесцвечиваются. КС Гр+ бактерий обладает большой жёсткостью, многослойной муреиновой сетью; сопутствующие компоненты: тейхоевые кислоты, полисахариды, полипептиды, белки. Стенки Гр- бактерий содержат однослойную муреиновую сеть; сопутствующие компоненты: липопротеиды и липополисахариды, образующие наружную липопротеиновую мембрану, содержащую белки-порины: перенос многих низкомолекулярных веществ. Между внешней липопротеидной мембраной бактериальной стенки и ПМ лежит периплазматическое пространство (периплазма 10 нм в толщину). Периплазма содержит тонкий (1-3 нм) муреиновый слой и раствор с гидролитическими ферментами и транспортными белками.
Под действием фермента лизоцима можно разорвать муреиновый каркас и растворить бактериальную стенку.
Протопласты бактерий нечувствительны к действию бактериофагов (вирусов, паразитирующих на бактериях).
КЛЕТОЧНЫЙ ЦИКЛ
Клеточный цикл - время существования клетки от деления до деления. Клетки с возрастом теряют способность к делению. А у некоторых, к примеру, аскарид, после прохождения эмбрионального развития рост организма происходит не за счёт роста числа клеток, а за счёт увеличения их размера. Есть клетки, полностью потерявшие способность к делению (клетки ЦНС). Смысл клеточного деления состоит в равномерном распределении редуплицированного ген. материала по двум новым клеткам.
Синтез ДНК происходит в интерфазе, в которой есть периоды, где синтеза ДНК не происходит (в них при авторадиографических исследованиях нет метки). Клеткам дали импульсную клетку (к примеру, меченый тритием предшественник ДНК - тимидин). Сначала появились делящиеся, но не содержащие метки клетки - клетки, закончившие синтез ДНК до начала эксперимента; затем появились меченые делящиеся - которые в момент мечения синтезировали ДНК (были в S -периоде); снова появились делящиеся немеченые - те, которые в момент мечения не вступили в S -период; снова появились меченые делящиеся - которые вступили в деление второй раз.
G1: 2с
S:2-4с
G2:4с
В интенсивно делящихся клетках содержание РНК во время интерфазы увеличивается по крайней мере в 2 раза. После деления в G1-фазу поступают клетки, содержащие вдвое меньше белков и РНК, чем исходная родительская клетка. Начинается рост клеток, в основном за счёт накопления клеточных белков, что определяется увеличением кол-ва РНК на клетку. В течение всего митоза (от поздней профазы до средней телофазы) синтез РНК подавлен.В S-фазе кол-во РНК увеличивается в соответствии с возрастанием кол-ва ДНК, достигает максимума в середине G2. Синтез белка во время митоза падает до 25%, а максимум достигает, как и РНК, в середине G2. G1 характеризуется ростом клетки и подготовкой к синтезу ДНК (док-во: амёба увеличивается вдвое и только потом приступает к делению; если в период роста у амёбы вырезать часть цитоплазмы, можно задержать деление: оно не наступит, пока амёба не достигнет нужных размеров). В течение G1 происходит синтез ферментов, необходимых для образования предшественников ДНК (например, нуклеотидфосфокиназ), ферментов метаболизма РНК и белка, повышается активность ферментов, участвующих в энергетическом обмене.
Длительность S-периода зависит от: 1. скорости репликации ДНК 2. числа и величины репликонов 3. числа включённых репликонов 4. общего числа ДНК. При коротком S-периоде в репликацию включено большее число репликонов.
Пареллельно синтезу ДНК в S-периоде идёт синтез гистонов и их миграция в ядро, где они связываются с ДНК.
Длительность G2 обычно меньше остальных периодов интерфазы, может вообще выпадать. В G2 продолжается синтез РНК и белков.
Таким образом, синтез макромолекул в интерфазе происходит с предварительным опережением.
Чем выше специализация клетки, тем ниже её способность делиться, как будто у неё есть выбор: размножаться или дифференцироваться.
Гетерокарионы - клетки с разнородными ядрами (разного происхождения): например, человека и курицы, опухоли человека и эритроцита. Естественные гетерокарионы: при воспалительных процессах в соединительной ткани появляются многоядерные "клетки инородных тел", возникшие, вероятно, в рез-те слияния соседних клеток.
ДЕЛЕНИЕ КЛЕТОК
Прокариотические клетки делятся без образования специальных сложных аппаратов путём прямого бинарного деления.
Репликация бактериальной ДНК идёт почти непрерывно в течение всего времени клеточного цикла. В случае экспоненциального роста бактериальной культуры её клетки всё время как бы находятся на стадии S-фазы. Это происходит до тех пор, пока в окр. среде достаточно пит. в-в для роста числа клеток в геометрической прогрессии. Затем рост числа клеток падает, часть из них погибает, а другие превращаются в споры. В этом состоянии бактерии могут находиться до тысячи лет.
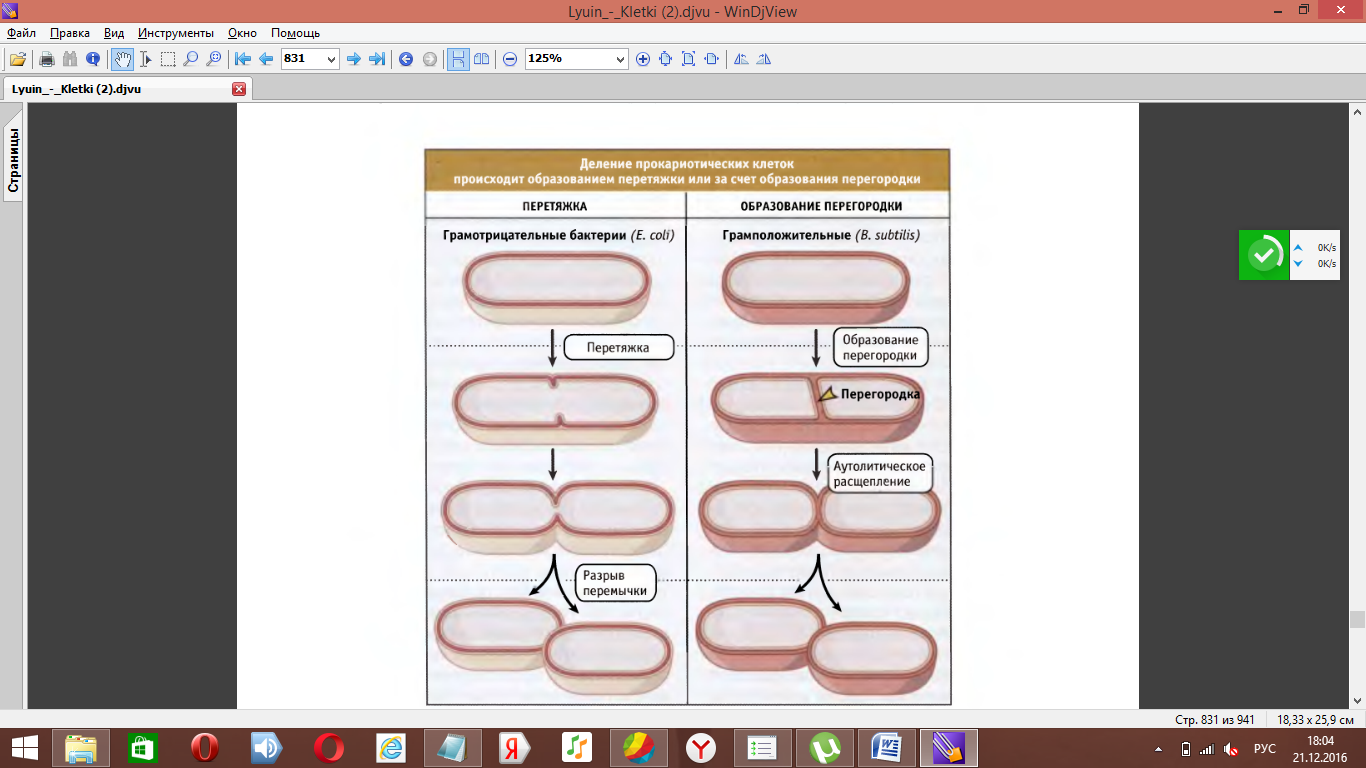
Деление прокариотической клетки контролируется главным образом на уровне образования кольца FtsZ (гомолог тубулина эукариот, белок, входящий в состав цитоскелета и формирующий мт) . Его образование может ингибировать MinCD.
Фактор блокирования нуклеоида: из-за своего объёма нуклеоид может предотвращать деление.

Эукариотические клетки могут делиться двояким способом: митоз (кариокинез, непрямое деление), амитоз (прямое деление).
Митоз
Этапы, не встречающиеся в интерфазе: интерфазные деконденсированные редуплицированные хромосомы переходят в конденсированную форму митотических хромосом, образуется специальный аппарат для сегрегации и переноса хромосом (ахроматиновый митотический аппарат), хромосомы расходятся к противоположным полюсам клетки, затем происходит деление тела клетки (цитотомия, цитокинез). Интерфаза по длительности во множество раз превосходит собственно митоз. Единственная фаза митоза, имеющая реальное начало, это анафаза - начало движения хромосом к полюсам; ещё анафаза самая короткая по времени фаза.
Фазы митоза:
Профаза
Первый видимый признак наступления митоза - появление в ядре конденсированных хромосом. У пойкилотермных животных крупные хромосомы, поэтому профаза длится несколько часов. У гомойотермных - небольшие, поэтому профаза длится менее 15 мин. Профаза обычно характеризуется появлением центросом (органелл в виде небольших точек, окружённых светлым участком), которые играют важную роль в образовании веретена: определяют его полюса, участвуют в нуклеации микротрубочек, из которых он образуется.
Клетки вступают в митоз при фосфорилировании одних белков и дефосфрилировании других. Эти процессы обеспечивают ферменты: киназы и фосфатазы. Очень важен киназный комплекс циклин B / CDK 1, при введении его в клетку индуцируется митоз. К концу профазы циклин B/CDK1 накапливается в ядре в неактивной форме. В клетку поступает cdc 25 фосфатаза, активирующая циклин B/CDK1. Активированный комплекс фосфорилирует многие ядерные белки, включая и те, которые обеспечивают целостность окружающей ядро оболочки. В рез-те эти белки теряют связь с ядерной мембраной, вызывают набухание ядра и разрыв окружающей его оболочки.
Разрыв ядерной оболочки знаменует собой начало прометафазы. В течение этой фазы хромосомы взаимодействуют с двумя центросомами и связанными с ними микротрубочками, образуя веретено. По мере присоединения хромосом к веретену они совершают конгрессию: хромосомы движутся по направлению к полюсам веретена и от них. Эти движения приводят к конгрегации (расхождению). Для большинства клеток прометафаза - самая продолжительная фаза. Продолжается, пока все хромосомы не расположатся на экваторе - наступает метафаза.
Сложная цепочка событий, приводящих клетку к метафазе, носит обратимый характер. Разрушение веретена в метафазе используется в экспериментальных целях для получения клеток, "заблокированных в метафазе".
Начало анафазы - две сестринские хроматиды каждой хромосомы начинают разделяться. Разделение хроматид в начале анафазы - ещё одна точка необратимости в митозе: она совпадает с разрушением "склеивающих" белков, скрепляющих хроматиды, и основной регуляторной киназы, вызывающей вступление клетки в митоз. Разделившиеся хроматиды расходятся к полюсам веретена. Анафаза А: расстояние между каждой хроматидой и полюсом, к которому она присоединена, сокращается. Анафаза В (в это же время): полюса веретена расходятся, растаскивая присоединённые к ним группы хромосом - элонгация веретена. По мере расхождения двух групп хромосом, веретено начинает разрушаться, между ними образуются межзональные структуры.
Телофаза (от греч. telo - конец) начинается, когда хромосомы формируют ядра у полюсов. Если при наступлении телофазы соседние анафазные хромосомы не соприкасаются, каждая из них образует собственное маленькое ядро. Эти ядра затем сливаются, образуя крупное ядро. Деление клетки на две: в плоскости, где в метафазе располагались хромосомы, образуется перетяжка, располагающаяся между двумя новыми ядрами и опоясывающая экваториальную область. Сжимаясь, перетяжка разделяет клетку на две примерно равные части (цитокинез). По мере сжатия перетяжки, межзональные структуры собираются вместе, образуя остаточное тельце - последнюю структуру, соединяющую две клетки. События телофазы требуют активации киназного комплекса циклин B/CDK1 и свидетельствуют о том, что клетка выходит из митоза.






