

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Оценка эффективности инструментов коммуникационной политики: Внешние коммуникации - обмен информацией между организацией и её внешней средой...
Генеалогическое древо Султанов Османской империи: Османские правители, вначале, будучи еще бейлербеями Анатолии, женились на дочерях византийских императоров...
Интересное:
Наиболее распространенные виды рака: Раковая опухоль — это самостоятельное новообразование, которое может возникнуть и от повышенного давления...
Что нужно делать при лейкемии: Прежде всего, необходимо выяснить, не страдаете ли вы каким-либо душевным недугом...
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Морфологию митотических хромосом лучше всего изучать в момент их наибольшей конденсации, в метафазе или в начале анафазы. Хромосомы животных и растений представляют собой палочковидные структуры разной длины с довольно постоянной толщиной. У большей части хромосом удаётся легко найти зону первичной перетяжки, которая делит хромосому на два плеча. Метацентрические хромосомы - равноплечие или почти равноплечие; субметацентрические - умеренно неравноплечие; акроцентрические - сильно неравноплечие с очень коротким, почти незаметным вторым плечом. В области первичной перетяжки расположена центромера. Это пластинчатая структура в форме диска, связанная тонкими фибриллами с телом хромосомы в области перетяжки. Центромера??? кинетохор??? является одним из центров полимеризации тубулинов (белков, из которых построены микротрубочки), от него отрастают пучки микротрубочек митотического веретена, идущие в направлении к центриолям. Эти пучки микротрубочек принимают участие в движении хромосом к полюсам клетки при митозе. Обычно хромосомы имеют только одну центромеру (моноцентрические), но бывают дицентрические и полицентрические.
В зоне первичной перетяжки присутствует ДНК (как и во всём теле хромосомы), элементарные фибриллы (что и в плечах хромосом). В зоне хромосомы, примыкающей к центромере, у многих видов локализована сателлитная ДНК (отличающаяся высоким уровнем повторённости нуклеотидных последовательностей).
Некоторые хромосомы имеют вторичную перетяжку, которая обычно расположена вблизи дистального (отдалённого от центра) конца хромосомы и отделяет маленький участок, спутник. Вторичные перетяжки также ??? (либо там расположены ЯО) ???называют ядрышковыми организаторами (ЯО), т.к. именно на этих участках хромосом в интерфазе происходит образование ядрышка. Здесь же локализована ДНК, ответственная за синтез рРНК. Эта ДНК относится к фракции, отличающейся умеренно повторяющимися нуклеотидными последовательностями.
|
|
Плечи хромосом оканчиваются теломерами, конечными участками, не способными соединяться с другими хромосомами или их фрагментами, в отличие от концов хромосом, лишённых теломерных участков в результате разрывов, которые могут присоединяться к таким же разорванным концам других хромосом.
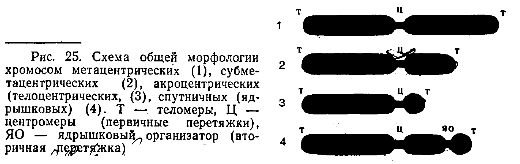
Размеры хромосом у разных организмов варьируют в широких пределах. Так, длина хромосом может колебаться от 0,2 до 50 мкм. Самые мелкие - у нек. простейших, грибов, водорослей. Наиболее длинные - у нек. прямокрылых насекомых, амфибий, лилейных. У человека в пределах 1,5-10 мкм. Число хромосом - видовой признак. Кариотип данного вида - совокупность числа, величины и морфологии хромосом.
Разработан ряд методов окрашивания (бэндинга), позволяющих выявить комплекс поперечных меток (полос, бэндов) на хромосоме. Каждая хромосома характеризуется специфическим комплексом полос. Гомологичные хромосомы окрашиваются идентично, за исключением полиморфных районов, где локализуются разные аллельные варианты генов. Аллельный полиморфизм характерен для многих генов и встречается в большинстве популяций.
Q -окрашивание. Первый метод дифференциального окрашивания хромосом был предложен Касперссоном, который показал, что при обработке препаратов митотических хромосом с помощью флуорохрома акрихин-иприта во флуоресцентном (люминесцирующем) микроскопе видна исчерченность по длине хромосом - участки с неодинаковой интенсивностью флуоресценции - Q -сегменты. Метод лучше всего подходит для исследования Y -хромосом и поэтому используется для быстрого определения ген. пола, выявления транслокаций (обменов участками) между X - и Y -хромосомами или между Y -хромосомой и аутосомами, а также для просмотра большого числа клеток, когда необходимо выяснить, имеется ли у больного с мозаицизмом по половым хромосомам (аномалией сочетания половых хромосом) клон клеток, несущих Y -хромосому.
|
|
G -окрашивание. После интенсивной предварительной обработки, часто с применением трипсина, хромосомы окрашивают красителем Гимзы. Под световым микроскопом на хромосомах видны светлые и тёмные полосы - G -сегменты. Хотя расположение Q -сегментов соответствует расположению G -сегментов, G -окрашивание оказалось более чувствительным и заняло место Q -окрашивания в качестве стандартного метода цитогенетического анализа. G -окрашивание даёт наилучшие результаты при выявлении небольших аберраций (отклонений от нормы, ошибок, нарушений, погрешностей) и маркерных хромосом (сегментированных иначе, чем нормальные гомологичные хромосомы).
R -окрашивание даёт картину, противоположную G -окрашиванию. Обычно используют краситель Гимзы или флуоресцентный краситель акридиновый оранжевый. Этим методом выявляют различия в окрашивании гомологичных G - или Q -негативных участков сестринских хроматид или гомологичных хромосом.
C -окрашивание используют для анализа центромерных районов хромосом (эти районы содержат конститутивный гетерохроматин) и вариабельной, ярко флуоресцирующей дистальной части Y -хромосомы.
T -окрашивание применяют для анализа теломерных районов хромосом. Эту методику, а также окрашивание районов ЯО азотнокислым серебром ( AgNOR -окрашивание) используют для уточнения результатов, полученных путём стандартного окрашивания хромосом.
ПРАКТИКУМ!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!
http://bookre.org/reader?file=480711&pg=77
Получение дифференциальной окраски хромосом связано, скорее всего, с различной способностью к искусственной деконденсации разных участков хромосом. Такая дифференциация позволила детально изучить строение хромосом человека. При обычных методах окраски весь набор из 46 хромосом человека принято подразделять по их размерам на 7 групп (A, B, C, D, E, F, G). Если при этом легко отличить крупные хромосомы от мелких, метацентрические от акроцентрических, то внутри групп трудно отличить одну хромосому от другой. Так, в группе С 6-я и 7-я хромосомы схожи между собой так же, как и с Х-хромосомой. Дифференциальное окрашивание позволяет чётко отличить эти хромосомы друг от друга. Этот приём цитологического анализа в сочетании с генетическими наблюдениями позволил начать составлять хромосомные карты человека, т.е. находить места расположения генов на определённых участках хромосом. Многие исследователи связывают способность отдельных участков хромосом к окрашиванию с их хим. различиями. Избирательно окрашивание связано с локализацией гетерохроматина.
|
|
ГЕТЕРОХРОМАТИН И ЭУХРОМАТИН
Термин "гетерохроматин" вначале был предложен для обозначения участков хромосом, интенсивно связывающихся с красителями (т.к. при переходе клетки от митоза к интерфазе определённые зоны различных хромосом или даже целые хромосомы остаются конденсированными - гетерохроматические). Гетерохроматические участки обычно располагаются в теломерных, центромерных, околоядрышковых районах хромосом, но могут входить и в состав их внутренних частей. Утеря даже значительных участков гетерохроматических районов хромосом не приводит к гибели клетки.
Эухроматические (эухроматин) - участки хромосом, которые в интерфазе деконденсируются. Считается, что эухроматические районы хромосом активны и содержат весь основной комплекс генов клетки или организма. В эухроматических районах хромосом реже наблюдаются разрывы при действии нек. хим. агентов (например, иприта).
Гетерохроматические районы оказывают сильное влияние на функции соседних с ними эухроматических районов, сближенных в рез-те хромосомных перестроек: эухроматические участки начинают конденсироваться, падает их функциональная активность, в них происходит инактивация генов.
Принято различать структурный (конститутивный) гетерохроматин и факультативный гетерохроматин. Факультативный гетерохроматин - участок хроматина, временно переходящий в конденсированное состояние, гетерохроматизируется. При этом значительно угнетается, а иногда и вовсе прекращается синтез РНК на таких участках хроматина. Но такое гетерохроматизированное состояние ядра может быть временным. При восстановлении функциональной активности ядер хроматин разрыхляется, переходит в эухроматическое состояние.
|
|
Структурный гетерохроматин такими переходами не обладает или почти не обладает. Функционально значение конститутивного гетерохроматина неясно. Предполагают, что он играет важную роль в поддержании общей структуры ядра, участвует в прикреплении хроматина к ядерной оболочке, может быть местом узнавания и ассоциации гомологичных хромосом при мейозе.
Хроматин интерфазных ядер представляет собой хромосомы, теряющие в это время свою компактную форму, разрыхляются, деконденсируются. Когда хромосома или её участок полностью деконденсированы, эти зоны называют диффузным хроматином. Диффузный хроматин соответствует эухроматину, который активен в синтетическом отношении. Процесс репликации и транскрипции обнаруживается именно в диффузном хроматине или на границе диффузного и конденсированного. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина (гетерохроматина). Конденсированный хроматин в животных клетках выстилает всю ядерную оболочку, прерываясь только в области пор. Глыбки конденсированного хроматина неправильной формы и различной величины разбросаны по всему ядру, некоторые локализованы возле ядрышка. В виде конденсированного хроматина в ядре существует конститутивный и факультативный гетерохроматин. Конститутивный гетерохроматин - истинный гетерохроматин, генетически неактивный, обогащён ДНК с высокоповторяющимися нуклеотидными последовательностями, локализован в теломерных, центромерных участках и вблизи района ядрышкового организатора. Факультативный гетерохроматин функционально неактивен в данном типе клеток. Чем более диффузен хроматин интерфазного ядра, тем выше в нём синтетические процессы. Падение синтеза РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина. Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде хромосом. В этот период хромосомы не несут никаких синтетических нагрузок. Итог: хромосомы могут находиться в двух структурно-функциональных состояниях: в рабочем (частично или полностью деконденсированном) и неактивном (в состоянии метаболического покоя при максимальной конденсированности).
|
|
|

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни...

Архитектура электронного правительства: Единая архитектура – это методологический подход при создании системы управления государства, который строится...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!