МП и (ПП):
МП (мембранный потенциал) – это разность потенциалов между наружной и внутренней поверхностями мембраны клетки в покое.
ПП (потенциал покоя) – та же разность потенц. Но уже в клетках возбудимой ткани.
Внутренняя сторона мембраны в норме всегда заряжена отрицательно по отношению к наружной.
Разность эл. Потенц. между мембранами, обусловлена неодинаковой конц. ионов, (в основном Na K Cl и различной её проницаемостью для этих ионов).
В среднем у клеток возбудимых тканей он достигает 50-80 мВ, (отриц внутри клетки). Обусловлен преимущественно ионами К.
3.Мембранный потенциал (потенциал покоя), механизм возникновения.
ПА возникает из-за 3 факторов:
1) ионная ассиметрия-
К- в клетке в 30-50 раз больше, чем снаружи
Na- вне клетки в 8-10 раз больше
Ca- больше вне
CI- вне клетки в 50 раз больше
2) Силективность мембраны (избирательность)-в покое мембрана пропускает только К.
3) Работы К-Na насоса - создаёт асимметрию и поддерживает постоянство потенциала.
Т.к. внутри клетки больше К и в покое мембраны открыты только калиевые каналы, К выходит ч/з мембрану наружу. Т.к. К связан силами электро-статического взаимодействия с органическими анионами (А), которые не могут пройти ч/з мембрану, то ионы К скапливаются снаружи, а А скапливаются внутри. !мембрана заряжается!
Постоянство ПП поддерживается К-Na насосами.
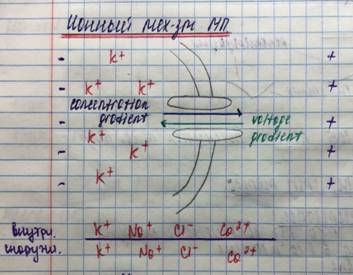 На мембране формируется К-равновесный потенциал, ибо уравновешиваются силы диффузии и электростатического взаимодействия.
На мембране формируется К-равновесный потенциал, ибо уравновешиваются силы диффузии и электростатического взаимодействия.
К - основной ион, обеспечивающий формирование МП(ПП), что подтверждается формулой Нернста:

К- Na насос: 1 цикл= 3Na наружу + 2К внутрь.
(4) Современные представления о процессе возбуждения. Потенциал действия и его фазы. Следовые явления.
(Будет очень хорошо если вы это вообще поймёте!)
ПД - это быстро-распостраняющийся по мембране клетки нервный сигнал, обеспечивающий передачу информации в центральной и периферической нервной клетке
ПД возникает в нервных, скелетных, гладкомышечных клетках, кардиомиоцитах, где обеспечивается связь возбуждения и сокращения
ПД возникает при действии раздражителя пороговой силы.
Условия возникновения ПД в клетке или ткани- деполяризация возбудимой мембраны до критического уровня.
Е0- исходный уровень ПП
Екр.- критический уровень деполяризации (КУД) 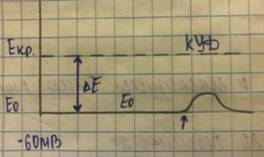 КУД- величина, до которой должен быть понижен ПП, чтобы возникло возбуждение.
КУД- величина, до которой должен быть понижен ПП, чтобы возникло возбуждение.
 Е- пороговый сдвиг мембранного потенциала, обозначаемый термином «порог деполяризации»
Е- пороговый сдвиг мембранного потенциала, обозначаемый термином «порог деполяризации»
Чем выше Е, тем большей силы необходим раздражитель для получения ПД  тем меньше возбудимость ткани.
тем меньше возбудимость ткани.
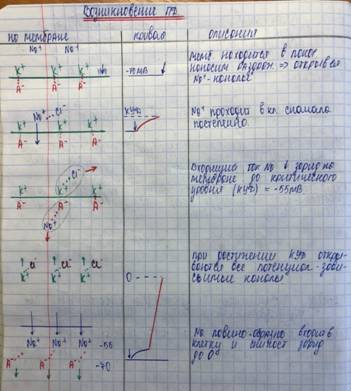
Деполяризация до КУД развивается медленно, при достижении МП критического уровня, проницаемость для Na резко возрастает.
Фазы ПД:
1) Деполяризация (восход.)
Na лавинообразно идёт в клетку (в 400-500 раз больше, чем в состоянии покоя)
Это ведёт к дальнейшему снижению МП, разность зарядов исчезает (МП=0)
Na продолжает проникать в клетку внутри  , а снаружи
, а снаружи  .
.
Перезарядка мембраны- реверсия/инверсия/овершут.
Спайк- пик потенциала
Когда на мембране устанавливается новое электрохимическое равновесие- проницаемость мембраны для Na резко угнетается.
Наступает инактивация натриевой проницаемости, что приводит к прекращению 1 фазы.
2) Реполяризация
Чтобы она произошла, надо убрать из клетки тот избыток , который образуется в связи с вхождение ионов Na.
Осуществляется 2-мя способами:
1. Увеличение проницаемости для К (в 10-15 раз) по сравнению с покоем.
2. Избыток Na удаляется работой Na-К насоса наружная сторона мембраны опять , выход К тормозится и становится равным исходному.
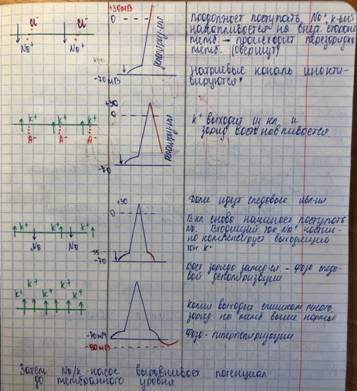
Следовыте явления (потенциалы):
1) (-) следовой потенциал = следовая деполяризация (СД)
СД- постепенное восстановление заряда на мембране, связанное с задержкой избытка Na внутри клетки, который не может быть быстро удалён работой Na-K насоса.
2) (+) следовой потенциал = следовая гиперполяризация (СГ)
СГ- связана с тем, что повышаемая проницаемость мембраны для К держится ещё некоторое время после восстановления исходного МП.
5.Сравнительная характеристика локального ответа и потенциала действия.
| ЛО
| ПД
|
| ЛО возникает в ответ на действие раздражителя подпороговой силы
| ПД возникает в ответ на действие раздражителя пороговой силы
|
| ЛО пропорционален силе подпоpогового раздражителя. ЛО зависит от силы раздражителя до тех пор, пока деполяризация, вызванная этим раздражителем, не достигнет своего критического уровня. В этот момент ЛО перестает быть ЛО, а превращается в ПД.
| ПД не зависит от силы раздражителя и подчиняется закону «все или ничего» - если раздражитель подпороговой силы, ПД нет, возникает только ЛО, если раздражитель пороговый, то возникает ПД и дальнейшее увеличение силы раздражителя не изменяет величину ПД, она стандартна для каждой клетки.
|
| ЛО может суммироваться до тех пор, пока изменения мембранного потенциала не достигнут КУД
| ПД не суммируется, потому, что ПД - это максимальный ответ, на который способна клетка.
|
| ЛО не передается по мембране, потому что при развитии ЛО не происходит пеpезаpядки мембраны (pевеpсии потенциала).
| ПД передается по мембране, потому, что при ПД происходит перезарядка мембраны, на короткое время она становится электроположитеьной. Следовательно, возникает разность потенциалов между возбужденным (+) и невозбужденным (-) участками нервного волокна.
|
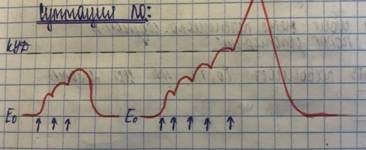
ЛО - если на возбудимую ткань наносить раздражение слабое, сост. 50-70% от пороговой силы раздражителя, то в ткани возникает местный нераспространяющийся процесс возбуждения- ЛО
Возникает частичная деполяризация за счёт увеличения проницаемости для Na.
Чем больше сила раздражения, тем в большей возрастает проницаемость мембраны для Na
 Свойства ЛО:
Свойства ЛО:
1) возникает при действии подпортгового раздражителя, не имеет четкого порога возникновения.
2) Подчинятся закону «силы» и не подчиняется закону «всё или ничего».
3) Не способен к распространению и локализуется в месте действия раздражителя на мембрану.
4) Возбудимость ткани во время ЛО повышается
5) ЛО могут суммироваться ( )
)
6.Особенности возбудимости нервных и мышечных волокон(?). Соотношение фаз возбудимости с фазами потенциала действия. Рефрактерность и ее причины.
Фазы ПД (Выше)
Фазы возбудимости:
ПЭ- первичная экзальтация (может ее не быть)- кратковременное повышение возбудимости. Если сейчас нанести подпороговый раздражитель, то возникнет ответная реакция.
I- ф. (абсолютной рефрактерности) = невозбудимости. Т.к. не реагирует ни на один из раздражителей.
II- ф. (относительной рефрактерности) - относительная невозбудимость. Ткань не может отвечать на сверхпороговые раздражения.
III- ф.(супернормальной возбудимости(вторичная экзальтация)) - возбудимость повышается ткань может отвечать на пороговые раздражёния.
IV-(субнормальная ф).- возбудимость снижена, ткань отвечает только на сверхпороговые раздражители
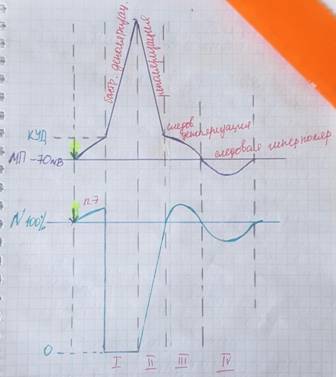 Сравнение фаз:
Сравнение фаз:
1) сверху ПД,
2) снизу возбуждение.
Рефрактерный период (период рефрактерности)- это период времени, после возникновения на возбудимой мембране потенциала действия, в ходе которого возбудимость мембраны снижается, а затем постепенно восстанавливается до исходного уровня.
1) Абсолютный рефрактерный период — интервал, в течение которого возбудимая ткань не способна генерировать повторный потенциал действия (ПД), каким бы сильным ни был инициирующий стимул.
2) Относительный рефрактерный период — интервал, в течение которого возбудимая ткань постепенно восстанавливает способность формировать потенциал действия. В ходе относительного рефрактерного периода стимул, более сильный, чем тот, который вызвал первый ПД, может привести к формированию повторного ПД.
Рефрактерный период обусловлен особенностями поведения потенциал-зависимых натриевых и потенциал-зависимых калиевых каналов возбудимой мембраны.
В ходе проведения потенциала действия потенциал-зависимые натриевые и калиевые ионные каналы переходят из одного состояния в другие. У натриевых каналов основных состояний три — закрытое, открытое и инактивированное. У калиевых каналов два основных состояния — закрытое и открытое.
При деполяризации мембраны во время проведения потенциала действия, натриевые каналы после открытого состояния (при котором и начинается ПД, формируемый входящим Na+ током) временно переходят в инактивированное состояние, а калиевые каналы открываются и остаются открытыми некоторое время после окончания ПД, создавая выходящий калиевый ток, приводящий мембранный потенциал к исходному уровню.
В результате инактивации натриевых каналов, возникает (1) абсолютный рефрактерный период. Позже, когда часть натриевых каналов уже вышла из инактивированного состояния, ПД может возникнуть. Однако для его возникновения требуются очень сильные стимулы, так как, во-первых, «рабочих» натриевых каналов всё ещё мало, а во-вторых, открытые калиевые каналы создают выходящий К+ ток и входящий натриевый ток должен его перекрыть, чтобы возник ПД — это (2) относительный рефрактерный период.
7.Законы раздражения возбудимых тканей (закон «силы», правило «все или ничего», закон «силы-времени», закон градиента раздражения (аккомодация)).
1) Закон силы - чем больше сила раздражителя, тем сильнее ответная реакция (до определённого предела).
Искл.: закон «всё или ничего»- до достижения пороговой величины ткань не отвечает (ничего), при достижении пороговой величины, ткань отвечает сразу максимально(всё). Дальнейшее увеличение силы раздражителя реакцию не меняет.
По этому закону работает сердечная мышца, отдельное нервное волокно, отдельное мышечное волокно, а также возник потенциал действия.

2) Закон «силы-времени»- соот. силы и длительности. Чем дольше мы действуем на ткань, тем сильнее ответная реакция до определённого предела.
Соотношение силы-длительности: чем больше силы раздражения, тем меньше времени нужно для получения ответной реакции.
Зависимость определяется формулой: I=a/t+b (гипербола)
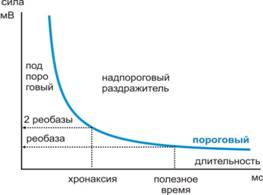 Реобаза- «порог силы тока»- min. сила тока, вызывающая возбуждение.
Реобаза- «порог силы тока»- min. сила тока, вызывающая возбуждение.
Порог времени (min. вр.)=1 мсек.
Хронаксия - min. время, необходимое для получения ответной реакции при силе тока =2 реобазы.
3) Закон градиента - если пороговая величина достигается очень медленно, то ткань успевает привыкать к действию раздражителя (происходит аккомодация) и поэтому на пороговую силу ткань не отвечает.
Таким образом важно значение скорости достижения пороговой величины.
8.Физиологические свойства скелетных и гладких мышц.
ФУНКЦИИ И СВОЙСТВА ГЛАДКИХ МЫШЦ
1. ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ. ГМ характеризуются нестабильным мембранным потенциалом (МП). Колебания МП независимо от нервных влияний вызывают нерегулярные сокращения, которые поддерживают мышцу в состоянии постоянного частичного сокращения — тонуса. МП ГМК не является отражением истинной величины потенциала покоя. При уменьшении МП мышца сокращается, при увеличении — расслабляется.
2. АВТОМАТИЯ. ПД ГМК имеют авторитмический характер, подобно потенциалам проводящей системы сердца. Это свидетельствует о том, что любые клетки гладких мышц способны к самопроизвольной автоматической активности. Автоматия гладких мышц, т.е. способность к а спонтанной деятельности, присуща многим внутренним органам и сосудам.
3. РЕАКЦИЯ НА РАСТЯЖЕНИЕ. В ответ на растяжение ГМ сокращается. Это вызвано тем, что растяжение уменьшает МП клеток, увеличивает частоту ПД и в конечном итоге — тонус ГМ. В организме человека это свойство гладкой мускулатуры служит одним из способов регуляции двигательной деятельности внутренних органов. Например, при наполнении желудка происходит растяжение его стенки. Увеличение тонуса стенки желудка в ответ на его растяжение способствует сохранению объема органа и лучшему контакту его стенок с поступившей пищей. В кровеносных сосудах растяжение, создаваемое колебаниями кровяного давления.
4. ПЛАСТИЧНОСТЬ. Изменчивость напряжения без закономерной связи с ее длиной. Так, если растянуть гладкую мышцу, то ее напряжение будет увеличиваться, однако если мышцу удерживать в состоянии удлинения, вызванным растяжением, то напряжение будет постепенно уменьшаться, иногда не только до уровня, существовавшего до растяжения, но и ниже этого уровня.
5. ХИМИЧЕСКАЯ ЧУВСТВИТЕЛЬНОСТЬ. ГМ обладают высокой чувствительностью к различным физиологически активным веществам: адреналину, норадреналину. Это обусловлено наличием специфических рецепторов мембраны гладкомышечных клеток..
ФУНКЦИИ И СВОЙСТВА СКЕЛЕТНЫХ МЫШЦ
Скелетная мышца обладает следующими важнейшими СВОЙСТВАМИ:
1) ВОЗБУДИМОСТЬЮ — способностью отвечать на действие раздражителя изменением ионной проводимости и мембранного потенциала.
2) ПРОВОДИМОСТЬЮ — способностью проводить потенциал действия вдоль и в глубь мышечного волокна по Т-системе;
3) СОКРАТИМОСТЬЮ — способностью укорачиваться или развивать напряжение при возбуждении;
4) ЭЛАСТИЧНОСТЬЮ — способностью развивать напряжение при растягивании.
Физиологической особенностью СЕРДЕЧНОЙ МЫШЦЫ является ее автоматизм. Возбуждение возникает периодически под влиянием процессов, протекающих в самой мышце. Способностью к автоматизму обладают определенные атипические мышечные участки миокарда, бедные миофибриллами и богатые саркоплазмой.
9.Виды сокращений скелетных мышц. Одиночное мышечное сокращение, его фазы. Тетаническое сокращение. Режимы мышечных сокращений.
У скелетной мышцы выделяют:
Одиночное сокращение
Оптимум и пессимум
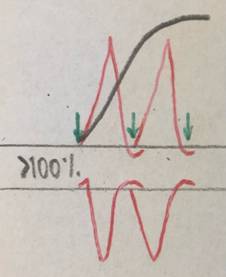
Если увеличивать частоту, то со временем каждый следующий импульс будет приходить в фазу повышенной возбудимости (супернормальной) ответная реакция ткани будет выше нормы. Это оптимум частоты.
Оптимум- такая частота тока, при которой амплитуда гладкого тетануса максимальна.
Пессимум – снижение амплитуды тетанического сокращения при увеличении частоты раздражений (выше оптимальной величины).
Пессимальная частота – максимальная частота (сверх оптимальной), при которой возникает минимальная амплитуда тетанического ответа.
| | | | |
Режимы мышечного сокращения:
1) Изотонический -сокращается мышца с изменением ее длины, без изменения тонуса
2) Изометрический -длина постоянная, тонус повышается
3) Смешанный (ауксометрический)- изменяется и длина, и тонус
10.Современная теория мышечного сокращения и расслабления.
Сокращение — это изменение механического состояния миофибриллярного аппарата мышечных волокон под влиянием нервных импульсов.
Теория скользящих нитей ( теория «скольжения»). В основе сокращения лежит взаимодействие между актиновыми и миозиновыми нитями миофибрилл вследствие образования поперечных мостиков между ними.
Саркомер — это участок миофибриллы между двумя соседними Z-линиями, структурно-функциональная единица поперечнополосатой мышечной ткани и других ее составляющих компонентов. К каждой Z-линии крепятся актиновые филаменты, а в центральной части саркомера располагаются миозиновые фибриллы.
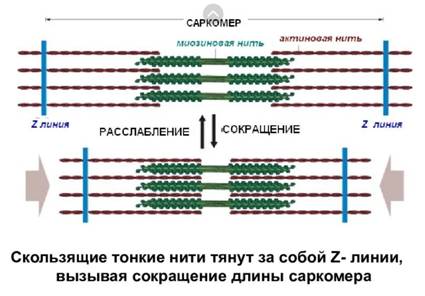 Во время скольжения сами актиновые и миозиновые нити не укорачиваются, но длина саркомера изменяется. В расслабленной, а тем более растянутой мышце активные нити располагаются дальше от центра саркомера, и длина саркомера больше. При изотоническом сокращении мышцы актиновые нити скользят по направлению к центру саркомера вдоль миозиновых нитей. Нити актина прикреплены к Z-мембране, тянут ее за собой, и саркомер укорачивается. Суммарное укорочение всех саркомеров вызывает укорочение миофибрилл, и мышца сокращается.
Во время скольжения сами актиновые и миозиновые нити не укорачиваются, но длина саркомера изменяется. В расслабленной, а тем более растянутой мышце активные нити располагаются дальше от центра саркомера, и длина саркомера больше. При изотоническом сокращении мышцы актиновые нити скользят по направлению к центру саркомера вдоль миозиновых нитей. Нити актина прикреплены к Z-мембране, тянут ее за собой, и саркомер укорачивается. Суммарное укорочение всех саркомеров вызывает укорочение миофибрилл, и мышца сокращается.
КЛАССИФИКАЦИЯ:
Морфологическая
1. Униполярные (имеют один отросток – аксон; характерны для ганглиев беспозвоночных);
2. Псевдоуниполярные (один отросток, делящийся на две ветви; характерно для ганглиев высших позвоночных).
3. Биполярные (есть аксон и дендрит, характерно для периферических и чувствительных нервов);
4. Мультиполярные (аксон и несколько дендритов – характерно для мозга позвоночных);
Функциональная
1.Афферентные (чувствительные, сенсорные – воспринимают сигналы из внешней или внутренней среды);
2.Вставочные, связывающие нейроны друг с другом (обеспечивают передачу информации внутри ЦНС: с афферентных нейронов на эфферентные).
3. Эфферентные (двигательные, мотонейроны – передают первые импульсы от нейрона к исполнительным органам).
4. Секреторные нейроны(нейросекреторные клетки) – это специализированные нейроны, по своей функции напоминающие эндокринные клетки. Они синтезируют и выделяют в кровь нейрогормоны, расположены в гипоталамической области головного мозга. Они регулируют деятельность гипофиза, а через него и многие периферические эндокринные железы.
Классификация:
1) По природе адекватного раздражителя (специфичности):
- механорецепторы - в коже, в сосудах, во внутренних органах, в опорнодвигательнмом аппарате слуховой и вестибулярной системы.
- терморецепторы (тепловые и холодовые) – в коже, в слизистых оболочках, во внутренних органах, в гипоталамусе, в среднем и продолговатом мозге.
- фоторецепторы – в сетчатке
- ноцицепторы (болевые) - в коже, в сосудах, во внутренних органах, в мышцах.
- хеморецепторы – (вкус, обоняние, изменение органического и газового состава крови О2/СО2) в каротидных и аортальных тельцах, в гипоталамусе, в продолговатом мозге.
- осморецепторы
- барорецепторы – (на изменение давления) в кровеносных сосудах
2) По дальности расположения:
- контактые
- дистантные
3) По порогу раздражителя:
- низкопороговые
- высокопороговые
4) По локализации раздражителя:
- экстрарецепторы
- интрарецепторы
- проприорецепторы
5) По механизму возникновения возбуждения:
- первичночувствующие
- вторичночувствующие
Свойства рецепторов
1) Специфичность. Большинство рецепторов приспособлены для восприятия только одного вида раздражителей. Специфичность таких мономодальных рецепторов не является абсолютной – практически любой рецептор реагирует на разные раздражители. Однако пороговая сила того раздражителя, к восприятию которого рецептор приспособлен, значительно ниже таковой для всех прочих раздражителей.
2) Чувствительность. Количественной мерой чувствительности сенсорного рецептора является абсолютный порог чувствительности – минимальная сила раздражителя, способная вызвать возбуждение рецептора.
Адаптацией называют явление ослабления возбуждения в рецепторе при действии длительного раздражителя постоянной силы.
Первичночувствующие рецепторы – воспринимают и трансформируют энергию раздражителя в нервный импульс непосредственно в чувствительном нейроне (в его отростке), и по его дендриту этот импульс напрямую передается к телу нейрона.
Пр.: тактильные, обонятельные, температурные, мышечные веретена.
Вторичночувствующие рецепторы – это специальные клетки, которые способны воспринимать раздражение и передавать его к окончанию афферентного волокна (дендрита) при помощи медиатора.
Пр.: зрительные, слуховые, вкусовые, вестибулярные
Общая особенность рецепторов – независимо от типа стимула и сложности строения анализатора, результатом действия раздражителя является изменение свойств клеточной мембраны, а именно – мембранного потенциала (МП) рецептора. Это изменение заряда получило название РЕЦЕПТОРНЫЙ ПОТЕНЦИАЛ (РП).
Механизм возникновения РП: в ответ на раздражение изменяется проницаемость мембраны для ионов Na. Входящий Na изменяет заряд мембраны и формирует РП.
Внесинаптическая область
4) синаптическая щель, величина которой составляет в среднем 50 нм
Пресинаптическая часть – специализированная часть терминали отростка нейрона, где расположены синаптические пузырьки и митохондрии. Пресинаптическая мембрана (плазмолемма) содержит потенциалозависимые Са2+-каналы. При деполяризации мембраны каналы открываются, и ионы Са2+ входят в терминаль, запуская в активных зонах экзоцитоз нейромедиатора.
Синаптические пузьрьки содержат нейромедиатор (ацетилхолин, аспартат, глутамат, ГАМК, глицин, адреналин, нейропептиды) Слияние синаптических пузырьков с пресинаптической мембраной происходит при увеличении концентрации Са2+ в цитозоле нервной терминали.
Активные зоны. – это участки утолщения мембраны, в которых происходит экзоцитоз. Активные зоны расположены против скоплений рецепторов в постсинаптической мембране, что уменьшает задержку в передаче сигнала, связанную с диффузией нейромедиатора в синаптической щели.
Постсинаптическая часть: Постсинаптическая мембрана содержит рецепторы нейромедиатора, ионные каналы.
МЕХАНИЗМ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ:
ПД достигает терминали. 1 терминаль даёт 1 аксону устанавливать контакт.
Ионы Са вызывают образование специального белкового комплекса, который включает в себя везикулу и структуры, расположенные около пресинапт. мембраны. Они связаны между собой белками экзоцитоза.
Излитию содержимого пузырька в щель способствует белок синаптопорин (формирует канал, по которому вбрасывается медиатор).
Квант медиатора- количество молекул, содержащееся в 1 везикуле. На 1 ПД выбрасывается 100 квантов АХ.
События в синапсе:
1) ПД приходит к терминалям аксона
2) Он деполяризует пресинаптическую мембрану
3) Са входит в терминаль, что приводит к выделению АХ
4) В синаптическую щель выделяется медиатор АХ
5) Он диффундирует в щель, связывается с рецепторами постсинаптической мембраны
6) Меняется проницаемость постсинаптической мембраны для Na
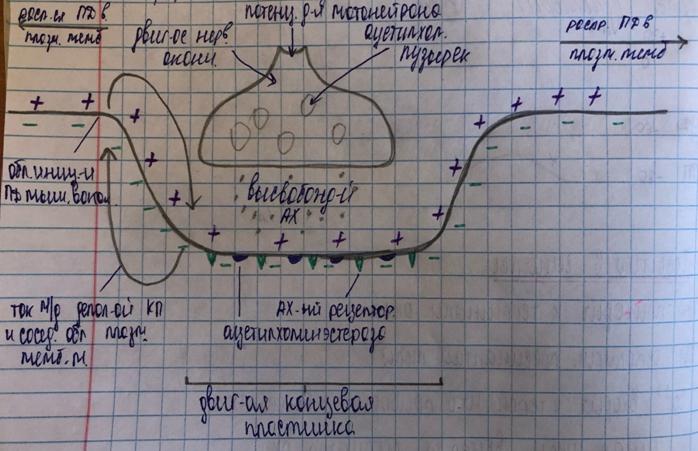
7) Na проникает в постсинаптическую мембрану, уменьшает её заряд – возникает потенциал концевой пластинки (ПКП)
На самой постсинаптической мембране ПД возникнуть не может, т.к. здесь нет потенциал-зависимых каналов, они хемозависимы.
8) ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и к распространению по мышечному волокну (5 м/с).
 Медиатором называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе.
Медиатором называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе.
Синтез медиаторов происходит в цитоплазме нейронов и нервных окончаний из компонентов внутри и внеклеточных жидкостей. В синапсах скелетной мыщцы вырабатывается только ацетилхолин. АХ - синтезируется из холина и уксусной кислоты с помощью фермента холинацетилазы.
-Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование.
Фракции медиатора:
1) доступная –рядом с пресинаптической мембраной.
2) депонированная – над 1-ой фракцией
3) диффузно рассеянная – наиболее удаленная от пресинаптической мембраны.
Судьба медиатора:
-связывание с рецептором
-разрушение ферментов (Ах-эстеразой)
-обратное поглощение в пресинапт. мембрану
-вымывание из щели и фагоцитоз.
16. Нервно-мышечный синапс, структурные элементы, механизм и особенности проведения возбуждения в синапсе.
Нервно-мышечный синапс – структура, которая обеспечивает передачу возбуждения с нервного волокна на мышечное. Состоит из
1) пресинаптической мембраны (мембрана терминали аксона мотонейрона СМ),
2) постсинаптической мембраны (мембрана мышечного волокна- двигательная пластинка)
3) синаптической щели между ними.
Пресинаптическая мембрана - покрывает поверхность синаптической бляшки (покрыв. концевую веточку аксона)
Постсинаптическая мемб.(концевая пластинка) - покрывает мышечное волокно в области синапса. Имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна повышение площади контакта. На ней находятся белковые структуры-рецепторы, способные связывать медиатор.
В синаптической щели есть мукополисахаридное плотное вещество в виде полосок, мостиков и содержит фермент-ацетилхолинэстераза.
В пресинаптических терминалях - большое количество пузырьков/везикул, заполненных медиатором.
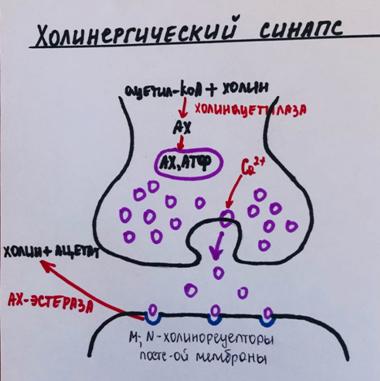
Механизм передачи возбуждения – химический. Химическое вещество, которое участвует в передаче возбуждения, называется медиатором. Медиатором в нервно-мышечном синапсе скелетных мышц является ацетилхолин. Ацетилхолин (АХ) находится в пресинаптическом нервном окончании в виде синаптических пузырьков (квантов).
ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ:
(1) возбуждение мембраны пресинаптического нервного окончания приводит к
(2) увеличению проницаемости пресинаптической мембраны для ионов Са (открываются потенциал-чувствительные кальциевые каналы),
(3) ионы кальция поступают из тканевой жидкости в нервное окончание.
(4) Они необходимы для выделения пузырьков медиатора (путем экзоцитоза).
(5) Медиатор (АХ) диффундирует к постсинаптической мембране и
(6) взаимодействует с холинорецепторами (белковыми молекулами, входящими в состав постсинаптической мембраны и имеющими высокое химическое сродство к ацетилхолину).
(7) В результате взаимодействия АХ с холинорецепторами открываются ионные каналы в постсинаптической мембране мышечного волокна. (Особенность ионных каналов постсинаптической мембраны: они хемо-чувствительные и проницаемы как для натрия, так и для калия).
(8) За счет движения ионов натрия в клетку и движения ионов калия из клетки происходит генерация постсинаптического потенциала – потенциала концевой пластинки (ПКП). ПКП имеет свойства локального ответа: зависит от количества медиатора, способен к суммации. Его амплитуда 30-70 мв.
(9) ПКП увеличивает возбудимость мембраны мышечного волокна (вызывает деполяризацию до критического уровня) и в околосинаптической зоне возникает ПД, который затем распространяется вдоль всего мышечного волокна.
(10) Ацетилхолин разрушается с помощью фермента ацетилхолинэстеразы (АХЭ) на холин и ацетат. Таким образом, холинорецепторы быстро освобождаются от медиатора. Холин возвращается в нервное окончание (с помощью специального активного транспорта) и используется для синтеза новых порций медиатора.
ОСОБЕННОСТИ ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ ЧЕРЕЗ ХИМИЧЕСКИЙ СИНАПС:
(1) одностороннее проведение (только от нервного волокна к мышечному волокну);
(2) синаптическая задержка (время, необходимое на выделение медиатора, диффузию его и т.д.)
(3) низкая лабильность (синапс способен проводить только 100 импульсов в сек)
(4) высокая утомляемость (связана с истощением запасов медиатора). В системе нерв–синапс–мышца утомление раньше всего развивается в синапсе вследствие более низкой скорости происходящих в нем химических процессов по сравнению с проведением электрических импульсов в нерве; уменьшения резерва медиатора и ионов кальция; снижения чувствительности рецепторов постсинаптической мембраны к действию медиатора.
(5) высокая чувствительность к действию химических блокаторов, которые связываются с холинорецепторами и нарушают нервно-мышечную передачу возбуждения. Связываясь с некоторыми из этих веществ, рецепторы постсинаптической мембраны образуют с ними более прочную связь, чем с естественным медиатором, но при этом ионные каналы не активируются и деполяризации постсинаптической мембраны не возникает. Рецепторы постсинаптической мембраны оказываются заблокированными "конкурентным" веществом, поэтому ни нервный импульс, ни выделяющийся в синаптическую щель медиатор не способны вызвать возбуждение и сокращение мышцы до тех пор, пока не кончится действие данного вещества.
(6) Суммация возбуждения соседних постсинаптических мембран
ФИЗИОЛОГИЯ КРОВИ
Состав крови
Цельная кровь состоит из жидкой части крови – плазмы – 54-64% (55 - 60%) и форменных элементов – эритроцитов, лейкоцитов и тромбоцитов - 36-46% (40 - 45%).
Количество крови в организме составляет 6-8% от массы тела, то есть у человека массой 65-70 кг количество крови 4,5-5 литров.
(Состав плазмы)
Плазма крови состоит из воды (90-92%) и сухого остатка (8 -10%).
Сухой остаток представлен:
Органическими соединениями.
Из них:
а) белки - 7-8%(Альбумины (до 50г/л), Глобулины (20-30г/л). Фибриноген (2-4 г/л);
б) остаточный азот в составе аммиака, мочевины, мочевой кислоты, креатина, креатинина 14-28 ммоль/л;
в) углеводы в виде глюкозы 3,6-6,5 ммоль/л;
г) липиды, в зависимости от характера принятой пищи – 2,0-4,0 г/л.
Показатель Гематокрита
Гематокрит – это доля объема крови, которую занимают форменные элементы. Этот показатель дает представление о процентном соотношении между объемом плазмы и объемом форменных элементов в крови.
В норме гематокрит составляет у женщин 36-42 (36-42%), у мужчин 40-48 (40-48%). Увеличение гематокрита наблюдается при эритроцитозе, уменьшение – при эритропении или микроцитозе (уменьшение размера эритроцитов) на фоне неизменной Гематокрит концентрации эритроцитов.
ИОНЫ КРОВИ
Основными ионами в плазме крови являются ионы натрия, калия, кальция, бикарбонатов, фосфатов и т.д. Они входят в состав ферментов, участвуют в процессах образования клеток крови и гемоглобина.И наверное ещё что-то они делают.
СОЛИ ПЛАЗМЫ КРОВИ
Минеральные соли в плазме крови представлены электролитами (катионы= натрия калия кальция и магния +анионы= хлориды + бикарбонаты) и они поддерживают осмотическое давление крови (в норме 7.3 атм).
Факторы влияющие на СОЭ
1) Количество и форма самих эритроцитов. При увеличении их количества и изменении формы эритроциты труднее склеиваются и медленнее оседают. И, наоборот, чем меньше эритроцитов, тем быстрее они оседают.
2) Белки плазмы крови. СОЭ снижается при увеличении содержания в плазме альбуминов и повышается при увеличении содержания фибриногена и глобулинов.
3) Холестерин, антигены, иммуноглобулины или антитела, образующиеся в избытке при патологических состояниях, липопротеины, мукополисахариды ускоряют СОЭ.
4) РН крови: ацидоз замедляет, а алкалоз ускоряет СОЭ.
5) Минеральные соли. Снижение солей кальция ускоряет СОЭ и, наоборот.
7.Буферные системы крови – стабилизируют рН крови
4 Разновидности
1. Гемоглобиновая. Она представлена восстановленным гемоглобином (ННb) и его калиевой солью (КНb). Это самая мощная буферная система крови, на ее долю приходится 75% буферной емкости крови.
2. Карбонатная, представлена – гидрокарбонатом натрия и угольной кислотой (NaHCO3/H2CO3)
3. Фосфатная буферная система. Включает соли фосфорной кислоты (Na2HPO4 и NaH2PO4).
4. Белковая буферная система, обусловлена наличием у белков плазмы амфотерных свойств.
8.Ацидоз – это сдвиг рН крови в кислую сторону за счёт увеличения количества кислых продуктов обмена.
9.Алкалоз – это сдвиг рН крови в щелочную сторону вследствие увеличения количества щелочей в крови.
Участие в иммуннитете
3)Детоксицирующая функция – способность адсорбировать токсические вещества и их инактивировать.
Виды гемолиза.
1. Химический – при действии на мембраны эритроцитов химических веществ (например, при отравлении): нитратов и нитритов, анилиновых красителей, кислот (соляная, уксусная), эфира, бензина, этилового спирта
2. Механический – возникает при сильных механических воздействиях на кровь (сильное встряхивание бутылки с кровью).
3. Термический – наблюдается при замораживании и размораживании крови. Разрушение оболочки эритроцитов при этом происходит кристалликами льда.
4. Биологический – развивается при переливании несовместимой крови, при укусах змей, насекомых, под влиянием гемолизинов.
5. Аутогемолиз – наблюдается при образовании в организме антител против собственных эритроцитов (при аутоиммунных заболеваниях).
6. Осмотический (коллоидно-осмотический) – возникает в гипотонической среде.
Строение
Является белком который относится к группе хромопротеидов (гемопротеидов), и





