1) вентиляция легких (внешнее дыхание) (обмен газов между атмосферным воздухом и альвеолами легких),
2) диффузия газов в легких (обмен газов в легких между альвеолярным воздухом и кровью),
3) транспорт газов кровью (процесс переноса кислорода от легких к тканям и углекислого газа от тканей к легким),
4) диффузия газов в тканях (обмен газов между кровью капилляров большого круга кровообращения и клетками тканей),
5) тканевое дыхание (потребление кислорода и выделение углекислого газа клетками организма) – биологическое окисление в митохондриях клетки.
Внешнее дыхание — это газообмен между организмом и окружающим его атмосферным воздухом. Осуществляется в два этапа:
1. обмен газов между атмосферным и альвеолярным воздухом
2. газообмен между кровью легочных капилляров и альвеолярным воздухом.
Аппарат внешнего дыхания – дыхательные пути, легкие, плевра, скелет грудной клетки и ее мышцы, диафрагма. Функция – обеспечение организма кислородом и освобождение его от избытка углекислого газа. О функциональном состоянии аппарата внешнего дыхания можно судить по ритму, глубине, частоте дыхания, по величине легочных объемов, по показателям поглощения кислорода и выделения углекислого газа и т. д.
2.Вентиляция лёгких. Биомеханика вдоха и выдоха.
Вентиляция легких осуществляется за счет создания разности давления между альвеолярным и атмосферным воздухом: при вдохе давление в альвеолярном пространстве значительно снижается и становится меньше атмосферного, поэтому воздух из атмосферы входит в легкие, смешиваясь с альвеолярным воздухом. При выдохе давление вновь меняется, т.е. давление в легких приближается к атмосферному или даже становится выше его, что приводит к удалению очередной порции воздуха из легких.
Биомеханика внешнего дыхания
Увеличение объема грудной полости обеспечивает вдох (инспирацию), уменьшение – выдох (экспирацию). Вдох и выдох составляют дыхательный цикл. Изменение объема грудной полости совершается за счет сокращений дыхательных мышц.
Акт вдоха (инспирация) – процесс активный. К инспираторным мышцам относятся диафрагма, наружные косые межреберные и межхрящевые. Главная мышца – диафрагма. При ее сокращении уплощается купол, органы каудально сжимаются, и стенка живота становится выпуклой. Одновременно диафрагма поднимает нижние ребра, что приводит к увеличению объема грудной полости не только в вертикальном направлении Сокращение наружных межреберных и межхрящевых мышц приводит к увеличению объема грудной полости в сагиттальном и фронтальном направлениях.При глубоком вдохе сокращаются также вспомогательные мышцы: лестничные, грудино-ключично-сосцевидная, большая и малая грудные, передняя зубчатая, а также трапециевидная, ромбовидная, поднимающая лопатку. Таким образом, в акте вдоха участвуют мышцы-разгибатели.
При спокойном дыхании выдох осуществляется пассивно, за счет расслабления мышц вдоха. При глубоком выдохе сокращаются экспираторные мышцы – мышцы брюшной стенки (косые, поперечная и прямая), внутренние косые межреберные мышцы, мышцы, сгибающие позвоночник. Такой выдох называется активным. Диафрагма и мышцы живота действуют вместе, когда должно быть создано высокое брюшное давление. Энергия мышц затрачивается на эластическое сопротивление грудной клетки, эластическое сопротивление легких, вязкое сопротивление перемещаемых тканей, аэродинамическое сопротивление дыхательных путей, тяжесть перемещаемой грудной клетки и верхних конечностей.
3.Отрицательное давление в плевральной полости, происхождение и роль в механизме внешнего дыхания, его изменения в разные фазы дыхательного цикла.
Внутриплевральное давление — давление в герметично замкнутой плевральной полости между висцеральными и париетальными листками плевры. В норме это давление является отрицательным относительно атмосферного. Внутриплевральное давление возникает и поддерживается в результате взаимодействия грудной клетки с тканью легких за счет их эластической тяги.Чем сильнее растягиваются легкие, тем выше становится их эластическая тяга и тем ниже падает давление в плевральной полости.
Величина отрицательного давления в плевральной полости равна:
v к концу спокойного вдоха – 5-7 мм рт.ст..
v к концу максимального вдоха – 15-20 мм рт.ст.,
v к концу спокойного выдоха – 2-3 мм рт.ст.,
v к концу максимального выдоха - 1-2 мм рт.ст.
Отрицательное давление в плевральной полости обусловлено так называемой эластической тягой легких – силой, с которой легкие постоянно стремятся уменьшить свой объем. Эластическая тяга легких обусловлена тремя факторами:
1. наличием в стенках альвеол большого количества эластических волокон;
2. тонусом бронхиальных мышц;
3. поверхностным натяжением пленки жидкости, покрывающей стенки альвеол.
Вещество, покрывающее внутреннюю поверхность альвеол, называется сурфактантом.
4.Диффузия газов в лёгких и тканях. Парциальное давление газов(кислород,углекислый газ) в альвеолярном воздухе,их напряжение в крови и тканях.
Диффузия газов в легких происходит вследствие разницы парциального давления газов в альвеолярном воздухе и в крови.
Парциальное давление кислорода в альвеолах (100 мм рт.ст.) значительно выше, чем напряжение кислорода в венозной крови, поступающей в капилляры легких (40 мм рт.ст.).
Градиент парциального давления углекислого газа направлен в обратную сторону (46 мм рт.ст. в начале легочных капилляров и 40 мм рт.ст. в альвеолах). Эти градиенты давлений являются движущей силой диффузии кислорода и углекислого газа в крови и в альвеолярном воздухе. Молекулы газа в силу диффузии переходят из области большего парциального давления в область низкого парциального давления.
Проницаемость мембраны для газа выражают величиной диффузионной способности легких – это количество газа, проникающего через легочную мембрану за 1 мин. при градиенте давления в 1 мм рт.ст.
В норме диффузионная способность для О2 составляет 20-25 мл/мин/мм рт.ст. Коэффициент диффузии для СО2 в 20-25 раз больше, чем для кислорода (диффузионная способность для СО2 составляет 600 мл на 1 мм рт.ст. в минуту).
При прочих равных условиях углекислый газ диффундирует через определенный слой среды в 20-25 раз быстрее, чем кислород. Именно поэтому обмен СО2 в легких происходит достаточно полно, несмотря на небольшой градиент парциального давления этого газа.
Диффузия газов в тканях.
1) Парциальное напряжение СО2 в тканях равно 60 мм рт. ст. парциальное напряжение О2 в тканях снижается до нуля.
2) В тканевой жидкости напряжение СО2 составляет 46 мм рт.ст., а кислорода 20 - 40 мм рт.ст.
3) В артериальной крови, притекающей к тканям напряжение СО2 – меньше 40 мм рт.ст., а напряжение О2 чуть более 100 мм рт.ст. Происходит газообмен, О2 переходит в тканевую жидкость и в ткани, СО2 – в кровь.
4) В результате кровь становится венозной и в ней напряжение СО2 равно 46 мм рт.ст., а напряжение О2 – 40 мм рт.ст.
Кислород поступивший в ткани по градиенту парциального давления из крови тканевых капилляров достигает митохондрий. Переходу кислорода в ткани способствует раскрытие дополнительных капилляров, под влиянием продуктов обмена веществ, в частности гистамина.
Тканевое дыхание – это механизмы доставки и потребления кислорода, а также образование и выделение СО2 из ткани. Путь кислорода охватывает три отрезка: транспорт кровью; диффузию из капилляров большого круга кровообращения в клетки и их митохондрии; химические реакции с цитохромной системой митохондрий для образования АТФ.
На обмен кислорода и углекислого газа в тканях влияют площадь обменной поверхности, количество эритроцитов, протекающих по капиллярам в единицу времени, величина диффузионного расстояния и коэффициенты диффузии тех сред, через которые осуществляется перенос газов.
Парциальное давление – это когда каждый газ в смеси оказывает давление в соответствии с его процентным содержанием и независимо от наличия других газов.

5.Транспорт кислорода кровью. Кривая диссоциации оксигемоглобина, её характеристика.
Транспорт кислорода.
Большая часть кислорода переносится кровью в виде химического соединения с гемоглобином, образуя оксигемоглобин. Присоединение кислорода к гемоглобину (оксигенация гемоглобина) происходит без изменения валентности железа.
Степень насыщения гемоглобина кислородом, т.е. образование оксигемоглобина, зависит от напряжения кислорода в крови. Графически эту зависимость отражает так называемая кривая диссоциации оксигемоглобина.
Кривая диссоциации оксигемоглобина - это график, отображающий зависимость от напряжения кислорода в крови скорости реакций связывания (ассоциации, стрелка вправо) кислорода гемоглобином в лёгких и высвобождения (диссоциации, стрелка влево) кислорода оксигемоглобином в тканях. Эта кривая имеет S – образную форму.
Крутая часть графика диссоциации оксигемоглобина соответствует напряжению кислорода в тканях. Здесь усиливается диссоциация HbО2, и О2 из крови переходит в ткани. Отлогая правая часть графика соответствует высоким напряжениям кислорода и свидетельствует о том, что в этих условиях содержание оксигемоглобина мало зависит от напряжения кислорода и его парциального давления в альвеолярном воздухе. Эта часть графика соответствует процессам образования оксигемоглобина в легких.
Сродство гемоглобина к кислороду изменяется в зависимости от многих факторов. Как правило, все эти факторы смещают кривую, увеличивая или уменьшая ее наклон, но не изменяя при этом ее S – образную форму.
Если сродство гемоглобина к кислороду повышается, то процесс идет в сторону образования оксигемоглобина, и график диссоциации смещается влево. Это наблюдается при снижении напряжения углекислого газа при понижении температуры, при сдвиге рН в щелочную сторону (рис.12).
При снижении сродства гемоглобина к кислороду процесс идет больше в сторону диссоциации оксигемоглобина, при этом график диссоциации смещается вправо. Это наблюдается при повышении парциального давления углекислого газа, при повышении температуры, при смещении рН в кислую сторону.
При ряде патологических состояний наблюдаются изменения процесса транспорта кислорода кровью. Ряд заболеваний (например, некоторые виды анемий) сопровождается сдвигами кривой диссоциации оксигемоглобина вправо (реже – влево).
6.Дыхательный центр. Современные представления о его структуре и локализации. Регуляторные влияния на дыхательный центр со стороны высших отделов головного мозга.
Дыхательный центр (ДЦ) – совокупность нейронов специфических (дыхательных) ядер продолговатого мозга, способных генерировать дыхательный ритм. В продолговатом мозге имеется 2 скопления дыхательных нейронов:
1. одно из них находится в дорсальной части, недалеко от одиночного ядра – дорсальная дыхательная группа (ДДГ);
2. другое расположено вентральнее, вблизи от двойного ядра – вентральная дыхательная группа (ВДГ), где локализованы центры вдоха и выдоха. В дорсальном ядре были обнаружены два класса нейронов: инспираторные нейроны типа Iα и Iβ. При акте вдоха возбуждаются оба класса этих нейронов, но выполняют разные задачи:
v инспираторные Iα-нейроны активируют α-мотонейроны диафрагмальной мышцы, и, одновременно, посылают сигналы к инспираторным нейронам вентрального дыхательного ядра, которые в свою очередь, возбуждают αмотонейроны скелетных дыхательных мышц;
v инспираторные Iβ-нейроны, возможно с помощью вставочных нейронов, запускают процесс торможения Iα-нейронов.
В вентральном ядре были обнаружены два типа нейронов – инспираторные (от них возбуждение идет к альфа-мотонейронам скелетной дыхательной мускулатуры) и экспираторные (активируют экспираторные скелетные мышцы). Среди них были выделены следующие виды нейронов:
1. «ранние» инспираторные – активны в начале фазы вдоха (инспирации);
2. «поздние» инспираторные –активны в конце вдоха;
3. «полные» инспираторные – активны в течение всего вдоха;
4. постинспираторные – максимальный разряд в начале выдоха;
5. экспираторные – активны во вторую фазу выдоха;
6. преинспираторные – активны перед вдохом. Они выключают активную экспирацию (выдох).
Нейроны экспираторного и инспираторного отделов дыхательного центра функционально неоднородны, контролируют разные фазы дыхательного цикла и работают ритмически.
7.Рефлекторная саморегуляция вдоха и выдоха. Схема ритмической смены дыхательных фаз.
Рефлекторная регуляция дыхания с механорецепторов
В дыхательных путях человека находятся следующие типы механорецепторов:
1. рецепторы растяжения легких (медленноадаптирующиеся);
2. ирритантные (быстроадаптирующиеся), рецепторы слизистой оболочки дыхательных путей;
3. J-рецепторы (юкстаальвеолярные рецепторы капилляров легких).
Рецепторы растяжения легких (РРЛ) находятся в гладких мышцах трахеи и бронхов. Они связаны с ДЦ крупными миелинизированными афферентными волокнами n.vagus. Адекватным раздражителем этих механорецепторов является растяжение стенок воздухоносных путей. Механорецепторы имеют разные пороги раздражения (низко – и высокопороговые). Рецепторы растяжения легких обладают медленной адаптацией, так как задержка дыхания мало изменяет их активность. Именно с этих рецепторов осуществляется рефлекс саморегуляции дыхания – рефлекс Геринга-Брейера. У наркотизированных животных раздувание легких вызывает рефлекторное торможение вдоха и наступление выдоха, а резкое уменьшение объема легких приводит к активации вдоха. При перерезке блуждающих нервов эти реакции исчезают. Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких.
В эпителиальных и субэпителиальном слоях всех воздухоносных путей, а также в области корней легких расположены так называемые ирритантные рецепторы, которые обладают свойствами механо- и хеморецепторов. Рецепторы возбуждаются при резких изменениях объема легких, а также при пневмотораксе, коллапсе, ателектазах, при действии на слизистую пылевых частиц, паров едких веществ и некоторых биологически активных веществ. Эти рецепторы обладают быстрой адаптацией. Так, например, возбуждение ирритантных рецепторов механическими раздражителями приводит к увеличению частоты дыхания, сужению бронхов; химическими – вызывает кашлевой рефлекс.
J-рецепторы (юкстаальвеолярные рецепторы капилляров легких) располагаются в интерстиции альвеол, в местах их контакта с капиллярами. Эти рецепторы особенно чувствительны к интерстициальному отеку, легочной венозной гипертензии, эмболии мелких сосудов легких, а также к действию ряда биологически активных веществ (никотин, гистамин). Стимуляция J-рецепторов вызывает вначале апноэ, затем поверхностное тахипноэ, гипотензию и брадикардию.
Ритмическая смена дыхательных фаз. Совокупность дыхательных нейронов продолговатого мозга и моста в последнее время принято называть центральным механизмом дыхания (ЦМД).
В основе современных представлений о функционировании ЦМД лежит представление Бредли (1975 г.) о наличии в мозге двух нейронных блоков, выполняющих определенную задачу:
1. генератора центральной инспираторной активности (ЦИА) - Iα.
2. механизма выключения инспирации – пул Iβ и ИТ нейронов.
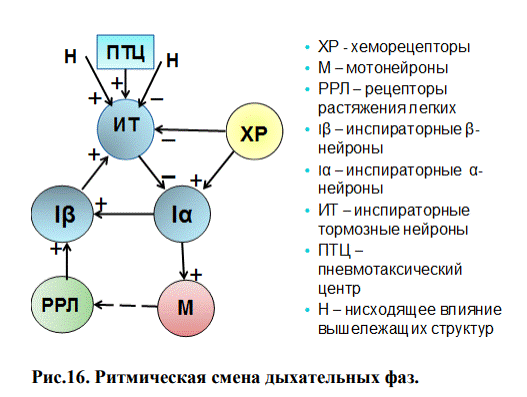
Генератор ЦИА представлен инспираторными нейронами типа Iα, локализованными в ДДГ продолговатого мозга. Они возбуждаются за счет потока афферентных импульсов с центральных и периферических хеморецепторов (ХР). Активность данных рецепторов находится в прямой зависимости от содержания в крови кислорода и углекислого газа (периферические рецепторы) и концентрации протонов в ликворе (центральные хеморецепторы).
При возбуждении нейронов типа Iα сигнал от них идет к ядрам дыхательных мышц спинного мозга (М), (рис.16). Одновременно от инспираторных нейронов типа Iα сигнал идет к инспираторным нейронам типа Iβ. Для активности нейронов типа Iβ важным является поток импульсов от рецепторов растяжения легких (РРЛ). Предполагают, что β – инспираторные нейроны возбуждают инспираторно-тормозящие нейроны (ИТ). Кроме того, на эти нейроны действует поток импульсов от нейронов, локализованных в варолиевом мосту, от так называемых пневмотаксического и апнейстического центров (ПТЦ). Они способствуют смене вдоха на выдох. ИТ нейроны возбудившись посылают тормозные импульсы к Iα, они тормозятся и перестают посылать импульсы к мотонейронам, наступает выдох. При этом Iβ не возбуждаются и не возбуждают ИТ нейроны, которые перестают тормозить Iα. Вновь наступает вдох.
8.Роль газового состава крови и регуляции дыхания (опыт Фредерика).Хеморецепторы, их локализация,роль.
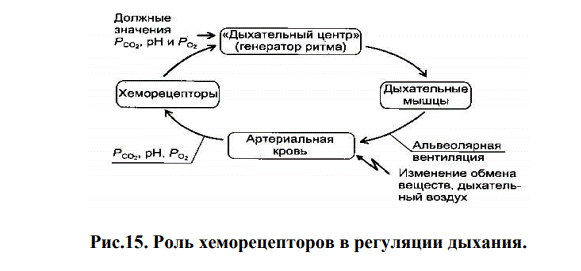
Регуляция внешнего дыхания направлена на поддержание постоянства газового состава внутренней среды организма. Значения рН артериальной крови и напряжения в ней СО2 и О2 не только непосредственно зависят от внешнего дыхания, но, в свою очередь, оказывают влияние на вентиляцию легких.
Опыт Фредерика – опыт, демонстрирующий значение СО2 и О2 в механизмах регуляции частоты и глубины дыхательных движений. Используются две подопытные собаки, имеющие перекрестное кровообращение головного мозга; если у одной собаки искусственно вызвать гипервентиляцию, то у второй разовьется апноэ вследствие вымывания из крови первой собаки СО2 и увеличения О2. Если у первой собаки пережать трахею, то у второй собаки будет наблюдаться учащение и углубление дыхательных движений (диспноэ), за счет крови, бедной О2, насыщенной СО2 и омывающей дыхательный центр у второй собаки.
Влияние СО2. Увеличение напряжения СО2 в артериальной крови (гиперкапния) приводит к повышению минутного объема дыхания. Как правило, при этом возрастают дыхательный объем и частота дыхательных движений.
У некоторых людей после длительного интенсивного дыхания (гипервентиляции) наблюдается временная остановка дыхания (апноэ) – эта остановка обусловлена отсутствием «физиологического стимулятора дыхания» - СО2, так как во время гипервентиляции СО2 вымывается и напряжение его в артериальной крови падает. Однако у ряда лиц после гипервентиляции возникает не апноэ, а лишь некоторое уменьшение минутного объема дыхания. Это позволяет предположить, что существует некий центральный механизм, запускающий дыхание даже в отсутствие стимулирующего действия СО2.
Влияние Н+. Если происходит снижение рН артериальной крови (ацидоз) по сравнению с нормальным уровнем, равным 7,36 вентиляция легких увеличивается. При возрастании рН выше нормы (алкалоз) вентиляция уменьшается, хотя и в несколько меньшей степени.
Влияние недостатка О2. Снижение напряжения О2 в артериальной крови сопровождается увеличением вентиляции легких. Это наблюдается при напряжении кислорода, равном 50-60 мм рт.ст. Артериальная гипоксемия может возникнуть во время пребывания на большой высоте, где атмосферное давление, а вследствие этого и парциальное давление О2 во вдыхаемом воздухе понижены. Напряжение О2 в артериальной крови снижается также при легочной патологии.
Роль хеморецепторов в регуляции дыхания. Контроль за нормальным содержанием во внутренней среде организма О2, СО2 и рН осуществляется периферическими и центральными хеморецепторами. Адекватным раздражителем для периферических хеморецепторов является уменьшение напряжение О2 артериальной крови, но в большей степени увеличение напряжение СО2 и уменьшение рН, а для центральных хеморецепторов – увеличение концентрации Н+ во внеклеточной жидкости мозга и напряжения СО2.
Периферические (артериальные) хеморецепторы находятся в основном в каротидных тельцах, расположенных в области бифуркации общих сонных артерий, и аортальных тельцах, находящихся в верхней и нижней частях дуги аорты. Сигналы от хеморецепторов аорты поступают по аортальной ветви блуждающего нерва, а от хеморецепторов каротидного синуса - по каротидной ветви языкоглоточного нерва (нерв Геринга) к дорсальной группе дыхательных нейронов продолговатого мозга. Более важную роль в возбуждении ДЦ играют хеморецепторы каротидного синуса.
Центральные (медуллярные) хеморецепторы чувствительны к изменению концентрации Н+ межклеточной мозговой жидкости. Они постоянно стимулируются Н+, концентрация которых зависит от напряжения СО2 в крови. При увеличении ионов Н+ и напряжения СО2 увеличивается активность нейронов ДЦ продолговатого мозга, растет вентиляция легких, и дыхание становится более глубоким. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы. Центральные хеморецепторы позднее реагируют на изменения газов крови, но возбудившись, обеспечивают прирост вентиляции на 60-80 %.
Отклонения, вызванные изменениями обмена веществ или состава дыхательного воздуха, приводят к изменению активности дыхательных мышц и альвеолярной вентиляции, возвращая значения напряжения О2, СО2 и рН к их должному уровню (приспособительная реакция).
пищеварение






