ТАБЛИЦА 2. ВОСПАЛИТЕЛЬНАЯ ПРИРОДА ЦИТОКИНОВ [1,2,102].
| Провоспалительные
| Интерлейкин-1β
|
| Интерлейкин-8
|
| Интерлейкин-12
|
| Интерлейкин-18
|
| Интерлейкин-23
|
| Фактор некроза опухолей-α (TNF-α)
|
| Моноцитарный Хемоаттрактантный Белок-1 (MCP1)
|
| Антивоспалительные
| Интерлейкин-10
|
| Трансформирующий ростовой фактор бета (TGF-β)
|
| Интерлейкин-4
|
| Интерлейкин-27
|
| Интерлейкин-35
|
| Переменные
| Интерферон-α *
|
| Интерлейкин-6 *
|
* Контрастные механизмы демонстрируют, что этот цитокин участвует как в ПРО-, так и в противовоспалительных процессах [2,4].
1. Arango Duque, G.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef]
2. Tilg, H.; Peschel, C. Interferon-alpha and its effects on the cytokine cascade: A pro- and anti-inflammatory cytokine. Leuk. Lymphoma 1996, 23, 55–60. [Google Scholar] [CrossRef]
3. Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
4. Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
ИНТЕРЛЕЙКИН 1
IL-1 представляет собой систему из трех ЦК: IL-1α, IL-1β, IL-1Ra (антагонист рецептора IL-1), и двух рецепторов R1 и R2. IL-1α и β кодируются разными (хотя и тесно сцепленными) генами и различаются по структуре и рI (α – 5,0; β – 7,0). Гомология их структуры составляет лишь 26%. Несмотря на незначительную гомологию, IL-1α и β конкурируют за один и тот же рецептор. Преобладающей формой IL-1 является IL-1β. Биологические свойства IL-1α и β очень сходны, либо идентичны. IL-1α активирует преимущественно Т-лимфоциты, обладает аутокринным и паракринным действием, в то время как IL-1β – многофункциональный ЦК с широким спектром действия, играет ключевую роль в развитии и регуляции неспецифической защиты и специфического иммунитета, один из первых включается в ответную защитную реакцию организма при действии патогенных факторов.
Основными продуцентами IL-1β являются макрофаги и моноциты. В синтезе данного ЦК также могут принимать участие лимфоциты, фибробласты. Клетки-мишени – иммунокомпетентные, эндотелиальные, эпителиальные клетки, фибробласты и др. IL-1β инициирует и регулирует воспалительные, иммунные процессы, активирует нейтрофилы, Т- и В-лимфоциты, стимулирует синтез белков острой фазы, ЦК (IL-2, -3, -6, TNF-α), молекул адгезии (Е-селектинов), прокоагулянтов, простагландинов. IL-1β повышает хемотаксис, фагоцитоз, гемопоэз, проницаемость сосудистой стенки, цитотоксическую и бактерицидную активность, оказывает пирогенный эффект и др.
IL-1 участвует в регуляции температуры тела, а его повышенная продукция приводит к развитию лихорадки. Известны факторы, снижающие биологическую активность IL-1. К ним прежде всего относят глюкокортикоиды и простагландины. Из экзогенных факторов следует указать циклоспорин А. В сыворотке крови лиц, которым был введен эндотоксин, в моче лихорадящих больных, а также в культуральной жидкости моноцитов, активированных in vitro, может быть обнаружен полипептид, специфически снижающий активность IL-1. Повышение уровня IL-1 наблюдается при различных воспалительных и АИЗ, включая септический шок, воспалительное поражение кишечника, РА, сахарный диабет 1 типа. Сильное повышение уровня IL-1 приводит к гипотензии, анорексии, разрушению хрящей в суставах.
Эндотелиальные клетки сосудов человека под влиянием IL-1α и β секретируют полипептиды, подобные тромбоцитарному фактору роста. Эти полипептиды могут стимулировать клеточную миграцию и пролиферацию и вызывать освобождение сосудистых медиаторов воспаления, что при значительном увеличении указанных ЦК может привести к диссеминированной внутрисосудистой коагуляции.
При множественных травмах в плазме наблюдается высокий уровень IL-1, -2, -6, и особенно резко увеличен уровень TNF. Отторжение почечного трансплантата сопровождается увеличением уровня IL-1, -6, TNF в плазме. Угроза прерывания беременности сопровождается увеличением продукции мононуклеарами периферической крови IL-1 и увеличением экспрессии рецептора IL-2 в субпопуляции Т-клеток. IL-1β принадлежит существенная роль в патогенезе СПИДа. При псориазе синтез IL-1α и β не снижается, но падает их функциональная активность. Низкая активность IL-1 может быть обусловлена генетически (возможно наследование аллеля гена IL-1, определяющего синтез IL-1 нормального содержания, но со слабой функциональной активностью). Повышенный уровень IL-1 отмечают при остром и хроническом миелоидном лейкозе. Увеличен уровень в плазме IL-1, -6, TNF при волосатоклеточном лейкозе. IL-1 стимулирует миелопоэз и ранние этапы эритропоэза (поздние – подавляет, будучи антагонистом эритропоэтина). IL-1β подавляет развитие В-лимфоцитов, участвует в выборе направления гемопоэза между миело- и В-лимфопоэзом (в пользу первого).
ИНТЕРЛЕЙКИН 2 (IL-2)
Этот ЦК с м.м. 15 кДа играет исключительно важную роль в реализации механизмов иммунного ответа. Продуцентами IL-2 являются Th1. Помимо участия IL-2 в дифференцировке и пролиферации Т-клеток, этот ЦК принимает непосредственное участие в реализации механизмов противоопухолевой защиты. Так, IL-2 повышает литическую активность NK-клеток, а также индуцирует клетки системы ЛАК (лимфокин-активированные киллеры). Кроме того, он усиливает секрецию IFNγ Т-лимфоцитами. Определение IL-2 является наилучшим показателем активации Т-клеток в in vitro тестах. Установлено, что IL-2 и IFNγ формируют эффекторные иммунологические механизмы, направленные на предотвращение пролиферации неотрансформированных клеток. У больных острым вирусным гепатитом в репликативный период регистрируется высокая спонтанная продукция IL-2.
ИНТЕРЛЕЙКИН 3 (IL-3)
IL-3 относится к семейству гемопоэтических ростовых факторов (м.м. 15-28 кДа). Клетками-продуцентами IL-3 являются Th1 и 2, а также ряд других клеток (В-лимфоциты, миелоидные и стромальные клетки костного мозга, кератиноциты). Активация гена IL-3 наблюдается через 4 ч после стимуляции клетки и поддерживается несколько суток. Секреция IL-3 подавляется циклоспорином А и глюкокортикоидами. IL-3 вместе с эритропоэтином поддерживает рост и дифференцировку клеток эритроидного ростка. В то же время IL-3 способен регулировать раннюю стадию дифференцировки В-лимфоцитов, поддерживает рост пре-В-клеток, а также усиливает секрецию IgG. IL-3, -4 и GM-CSF являются ростовыми факторами для тучных клеток. IL-3 усиливает продукцию гистамина клетками гемопоэтическойсистемы, но не влияет на активность гистамина в культуре клеток периферической крови взрослых доноров. IL-3 и GM-CSF вызывают формирование гранул эозинофилов.
ИНТЕРЛЕЙКИН 4 (IL-4)
Этот ЦК (м.м. 15-20 кДа) продуцируется Th2 и является фактором дифференцировки для Т- и В-клеток. Наиболее сильный эффект IL-4 оказывает на регуляцию образования других ЦК посредством участия в многочисленных биологических процессах, таких как иммунный ответ и воспалительные реакции. IL-4 ограничивает синтез макрофагами провоспалительных IL-1β, -6, -8, -12, TNF-α, образование высокоактивных метаболитов кислорода, азота. Кроме того, IL-4 служит кофактором пролиферации покоящихся В-лимфоцитов, а также индуцирует в этих клетках синтез IgE и IgG4. Известна способность IL-4 генерировать активность ЛАК и усиливать противоопухолевую активность макрофагов. Дисрегуляция секреции IL-4 является ключевой в развитии аллергопатологии. Показано, что МНК больных атопическими заболеваниями имеют усиленный ответ на рекомбинантный IL-4 по сравнению с ответом МНК здоровых доноров. Увеличение синтеза IgE в ответ на стимуляцию IL-4 приводит к усилению IgE-стимулированного синтеза ЦК тучными клетками, способными вырабатывать IL-4-6. При синдроме Сезари увеличено содержание IL-4. Уровень ЦК также заметно повышается у больных хроническим вирусным гепатитом С. В периоды обострения его количество увеличивается почти в 3 раза по сравнению с нормой, а во время ремиссии уровень IL-4 снижается, особенно на фоне проводимого лечения рекомбинантным IL-2.
ИНТЕРЛЕЙКИН 5 (IL-5)
Этот димерный белок с м.м. 50-60 кДа продуцируется Th2. IL-5 усиливает пролиферацию активированных В-клеток, а также экспрессию на них рецептора для IL-2 и синтез IgA. В нестимулированных В-клетках IL-5 индуцирует секрецию IgM и G. IL-5 является хемоаттрактантом для эозинофилов, вызывает их дегрануляцию при паразитарных инвазиях, играет роль в патогенезе аллергического воспаления, атопии. Противоопухолевая активность IL-5 связана со способностью участвовать в апоптозе.
ИНТЕРЛЕЙКИН 6 (IL-6)
Этот мономер с м.м. 19-34 кДа является фактором дифференцировки В-лимфоцитов в АТ-продуцирующие клетки. IL-6 индуцирует синтез белков острой фазы, в связи с чем (также как и IL-1 и TNF) может быть отнесен к ЦК воспаления. Показано, что IL-6 вызывает значительное повышение уровня мРНК с-sis гена (β-цепь РФТ) в культивируемых эндотелиальныхклетках человека, что может опосредовать воспалительные сосудистые эффекты. Повышение уровня IL-6наблюдается при многих патологических состояниях,в том числе при АИЗ, сердечной микседеме, РА, болезни Кастлемана, псориазе, мезангиопролиферативномгломерулонефрите, саркоме Капоши, алкогольномциррозе, лимфоме, миеломе и карциноме почек. УВИЧ-инфицированных лиц В-лимфоциты продуцируют увеличенное количество TNFα и IL-6. Есть данные обобнаружении повышенного уровня TNF-α и IL-6 в плазме крови при различных атопических реакциях, такихкак аллергия и астма. Данный ЦК регулирует пролиферацию эпителиальных клеток желчных протоков,клеток печени, образование гранулем, формированиефиброза при циррозе печени. Повышение концентрации IL-6 отмечено при обострениях язвенной болезни,панкреатита, глютеновой энтеропатии, болезни Крона,неспецифического язвенного колита, вирусного гепатита, первичного билиарного цирроза.
ИНТЕРЛЕЙКИН 7 (IL-7)
IL-7 – ЦК, стимулирующий гемопоэз. Является полипептидом с м.м. 20-40 кДа; продуцируется фибробластами и стромальными костномозговыми клетками. IL-7 стимулирует пролиферацию, но не дифференцировку пре- и про-В-клеток и не обладает активностью в отношении дифференцированных В-клеток. Также IL-7 стимулируется пролиферация незрелых и дифференцированных активированных Т-клеток. Он эффективен и в иммунотерапевтическом разрушении опухолевых клеток CD4+ Т-клетками. Совместно с IL-2 он может применяться в консолидативной иммунотерапии злокачественных новообразований у пациентов после трансплантации костного мозга. IL-7 может индуцировать апоптоз опухолевых клеток, вызывает дифференцировку клеток подгруппы острого миелобластного лейкоза.
ИНТЕРЛЕЙКИН 8 (IL-8)
IL-8 – низкомолекулярный ЦК воспаления; принадлежит к семейству хемокинов. Продуцируется под воздействием бактериальных эндотоксинов и ЦК, главным образом TNF и IL-1. Образуясь из общего для различных хемокинов предшественника, состоящего из 99 аминокислотных остатков (АКО), IL-8 содержит 72 АКО и существует в растворе в виде димера. Он известен как NAP-1 (активирующий нейтрофилы пептид-1), NAF (фактор активации нейтрофилов), GCF (хемотактильный фактор гранулоцитов) и NCF (хемотактильный фактор нейтрофилов). Активирует нейтрофилы, в меньшей мере другие гранулярные лейкоциты, вызывает их хемотаксис в очаг воспаления. Точно такой же эффект оказывается IL-8 на моноциты. Повышенный уровень IL-8 ассоциируется с хроническими и острыми воспалительными состояниями и коррелирует с тканевой инфильтрацией нейтрофилов при РА, с язвенным колитом. IL-8, появляясь после IL-1 и TNF в местах воспаления, играет важную роль при псориазе.
ИНТЕРЛЕЙКИН 10 (IL-10)
Природа иммунного ответа определяется цитокинами, которые продуцируются активированными клетками иммунной системы. При повреждении тканей цитокины вызывают цепь событий, ведущих к миграции нейтрофилов из кровеносного русла и формированию острого воспаления. В основе механизма их действия лежит способность влиять на дифференцировку, пролиферацию и гибель клеток [39].
Интерлейкин 10 - противовоспалительный цитокин, продукт гена человека IL10 [40]. Передача сигнала, опосредуемого интерлейкином 10, обеспечивается рецепторным комплексом, состоящим из 2 IL10RA (альфа-субъединиц) и 2 IL10RB (бета-субъединиц) [41]. IL-10 активирует STAT3-опосредуемый сигнальный путь. Фосфорилирование рецепторов проходит под действием киназ JAK1 и Tyk2 для альфа- и бета-субъединиц, соотв [41].
Интерлекин 10 – 35–40 кДа, продуцируется Т-хелперами 1-го и 2-го типа (Th-1 и Th-2), моноцитами, макрофагами и цитотоксическими клетками и имеет широкий спектр действия с выраженным иммуносупрессивным эффектом: подавляет пролиферацию и активность Т-клеток, продукцию синтеза ряда цитокинов, снижает активность макрофагов и моноцитов. IL-10 снижает активность Th-1 в большей степени, чем Th-2. IL-10 может стимулировать синтез IgE. В своем ингибирующем действии на клеточный иммунитет IL-10 синергичен с IL-4. IL-10 является родоначальником семейства цитокинов, в которое входят IL-19, IL-20, IL-22, IL-24 и IL-26.
Противовоспалительная активность IL-10 проявляется способностью снижать продукцию провоспалительных цитокинов, усиливать продукцию антагониста рецептора IL-1 и уменьшать адгезию лейкоцитов к эндотелиальным клеткам, активированным IL-1 (прим.:IL-1 – полипептид (15 - 17 кДа), описан как фактор, производимый мононуклеарными фагоцитами и усиливающий Т-клеточный ответ на антигены или поликлональные активаторы, т.е. костимулятор Т-клеточной активации. Считается, что IL-1 является лидирующим среди цитокинов воспаления).
Основной эффект IL-10 заключается в подавлении синтеза цитокинов Thl-клетками (т.е. он обладает действием, противоположным влиянию интерферона-гамма) и в снижении активности макрофагов, в том числе продукции воспалительных цитокинов. IL-10 подавляет экспрессию молекул МНС II класса, пролиферацию Т-клеток, вызванную митогенами, а также развитие гиперчувствительности замедленного типа. В то же время IL-10 выступает в качестве кофактора IL-2 и IL-7 в отношении пролиферации тимоцитов, служит синергистом IL-4, усиливает пролиферацию В-клеток, защищает их от апоптоза, повышает синтез IgM и IgA. В результате IL-10 способствует развитию гуморальной составляющей иммунного ответа, обусловливая антипаразитарную защиту и аллергическую реактивность организма.
Есть данные об исследовании полезной роли IL-10 при меланоме, карциноме, лимфоме, системной красной волчанке, аллергическом контактном дераматите, рассеянном склерозе, астме, а также по применению в клинической практике при ревматоидном артрите, псориазе, болезни Крона, гепатите С [42-44]. Таким образом, IL-10 служит важнейшим регулятором иммунного ответа, подавляющим активность макрофагов и Th1-клеток и обеспечивающим реализацию некоторых биологических эффектов Тh2. Будучи супрессивным цитокином, IL-10 обладает способностью подавлять иммунное воспаление и является важнейщим регулятором цитокинов, во многом определяющим направленность иммунных реакций.
P.S. Эффект IL-10 в отношении опухолевого роста не столь однозначен. Например, у IL-10-трансгенных мышей опухоли более агрессивны, а введение лимфоцитов от этих мышей способствует ускорению роста опухоли. Однако было показано ингибирующее действие IL-10-продуцирующих опухолевых клеток человека у иммуносупрессивных или бестимусных мышей. Таким образом, предполагается, что эффект зависит от состояния иммунокомпетентных клеток. Полученные данные о способности IL-10 влиять на антигенпрезентирующую способность дендритных клеток могут быть использованы для повышения эффективности противоопухолевых вакцин [45].
ИНТЕРЛЕЙКИН 11 (IL-11)
IL-11 синтезируется стромальными клетками костного мозга. Клетки-мишени – гемопоэтические предшественники остеокластов. Аминокислотная последовательность IL-11 человека и приматов идентична на 94%, а человеческого и мышиного – на 88%. IL-11 действует синергически с IL-3, -4, -7, -12, -13, SCF, Flt-3 лигандом и GM-CSF, стимулируя пролиферацию гемопоэтических клеток-предшественников. Он стимулирует мегакариоцитопоэз и тромбоцитопоэз совместно с IL-3, тромбопоэтином и SCF; эритропоэз, миелопоэз и лимфопоэз, модулирует гемопоэтическое микроокружение. Альвеолярные и бронхиальные эпителиальные клетки продуцируют IL-11 в ответ на воспалительные ЦК. IL-11 регулирует рост GI клеток эпителия. Он стимулирует развитие остеокластов, и ингибирует липогенез. Подобно IL-1 и -6, принимает участие в индукции синтеза белков острой фазы.
ИНТЕРЛЕЙКИН 12 (IL-12)
IL-12 является гликопротеином с м.м. 70 кДa, который состоит из двух гликозилированных субъединиц: р40 и р35, связанных между собой дисульфидными мостиками. Дисульфидные связи играют важную роль в биологической активности IL-12. Субъединицы кодируются двумя различными, независимыми друг от друга генами. Помимо обладающего биологической активностью р70-гетеродимера, клетки, продуцирующие IL-12, секретируют в большом количестве субъединицу р40, которая не является биологически активной. р40 секретируется с большим избытком по сравнению с биологически активным гетеродимером. Она участвует в связывании с рецептором, но р35 необходима для трансдукции сигнала. IL-12 секретируется, прежде всего, активированными макрофагами и влияет на иммунные клеточные реакции. IL-12 повышает литическую активность системы ЛАК. IL-12 действует как ростовый фактор при активации Т- и NK-клеток. При этом он действует в качестве индуктора секреции IFN-γ и ингибитора синтеза IgE, индуцированного IL-4. IL-12 активирует и цитотоксичность макрофагов, а дефицит его продукции макрофагами может значительно снижать противоопухолевую активность. IL-12 оказывает противоопухолевый эффект при раке легкого. Усиление роста опухоли, в частности, рака прямой кишки, ассоциируется со снижением продукции IL-12 и усилением продукции IL-10. Важным свойством IL-12 является усиление экспрессии FasL и индукция апоптоза. IL-12 ингибирует ангиогенез. Антиангиогенное действие IL-12 реализуется на уровне рецепторов протеинкиназ, адгезивных молекул, интегринов и других поверхностных структур, усиления продукции IFN-γ. В последние годы установлено, что IL-12 является ключевым ЦК в развитии Th1. Было показано, что Th1 способствуют патогенезу различных органоспецифических заболеваний с иммуновмешательством (аллергический энцефаломиелит, инсулин-зависимый диабет). IL-12 играет основную роль при АИЗ, резистентности к бактериальной или паразитической инфекции, антивирусном ответе, включая ВИЧ. Было показано, что IL-12 является мощным адьювантом при вакцинации.
ИНТЕРЛЕЙКИН 13 (IL-13)
IL-13 является белком, который продуцируется преимущественно в негликозилированной форме (10 кДа) активированными Т-клетками и мастоцитами. Функции IL-13 подобны биологической активности IL-4. Он является мощным модулятором активности моноцитов и В-клеток, но, в отличие от IL-4 и IL-13, не имеет прямого биологического влияния на Т-клетки. IL-13 оказывает ингибирующий эффект на продукцию других ЦК, стимулирующих начало воспалительного процесса при сепсисе или РА, причем в отличие от IL-4 его концентрация не снижается. IL-13 совместно с IL-4 и IL-10 принимает участие в иммунных реакциях Th2. У В-клеток он стимулирует секрецию IgG4 и IgЕ.
ИНТЕРЛЕЙКИН 15 (IL-15)
IL-15 – это недавно открытый ЦК, который обладает многими биологическими свойствами IL-2. IL-15 был открыт как ЦК (IL-T), продуцируемый линией Т-клеточного лейкоза взрослого человека (HuT-102) и стимулировавший пролиферацию Т-клеток. Высокоаффинные IL-15R были выявлены на различных клетках, включая T-, B- и NK-клетки, а также клетки нелимфоидного ряда. IL-15R состоит из трех субъединиц; специфической к IL-15Rα-, β- и γ-цепи, аналогичных рецепторной системе IL-2. Показано, что IL-15R стимулирует рост NK-клеток, активированных периферических Т-клеток, лимфоцитов, инфильтрирующих опухоль (TILs), и B-клеток. Показано, что IL-15 является хемоаттрактантом для Т-лимфоцитов человека и индуцирует активность ЛАК и NK и образование цитолитических эффекторных клеток. По-видимому, IL-15 также оказывает влияние на клетки, не связанные с иммунным ответом. Клетки скелетных мышц экспрессируют мРНК IL-15 и IL-15 R. Содержание IL-15 увеличивается при воспалительных заболеваниях желудка, тонкой и толстой кишки.
ИНТЕРЛЕЙКИН 16 (IL-16)
IL-16 (лимфоцитарный хемотаксический фактор – LCF) – это провоспалительный цитокин, хемоаттрактант CD4+ T-лимфоцитов, моноцитов и эозинофилов. Он может усиливать экспрессию IL-2R и HLA-DR, ингибировать T-клеточный рецептор (TcR)/CD3-зависимую активацию и способствовать подавлению транскрипции HIV-1. IL-16 – уникальный регулятор, не обладающий значительной гомологией с какими-либо другими охарактеризованными ЦК. Впервые IL-16 был идентифицирован как гомотетрамер, состоящий из отдельных мономеров с м.м. 14 кДа (130 АКО). У приматов его последовательность более чем на 90%. CD4 служит рецептором сигнальной трансдукции IL-16, поэтому данный ЦК способен взаимодействовать с СD4. Область молекулы CD4, с которой связывается IL-16, расположена внутри домена D4, перекрывая структуры, участвующие в формировании димеров CD4.
Источниками IL-16 служат эндотелиальные и тучные клетки, лимфоциты, макрофаги, синовиальные фибробласты и эозинофилы. IL-16 может секретироваться активированными CD8+ клетками в ответ на гистамин или серотонин. Экспрессия IL-16 связана с воспалительными процессами при астме, РА, СКВ, колитах, атопическом дерматите и рассеянном склерозе. Например, продукция IL-16 прямо пропорциональна количеству инфильтрирующих CD4+ T-клеток в эпителии больных астмой. У пациентов с III-IV стадией рака молочной железы, кишечника, почки, мочевого пузыря, матки, яичника в сыворотке крови обнаруживают повышенный уровень IL-16.
ИНТЕРЛЕЙКИН 17 (IL-17)
IL-17, впервые описанный как мышиный цитотоксический T-лимфоцит-ассоциированный антиген-8 (CTLA-8), продуцируется в первую очередь активированными Т-клетками памяти. Основываясь на исследованиях клеточных линий, предполагается, что IL-17R экспрессируется на тучных клетках, фибробластах, эмбриональных гепатоцитах, пре-B-клетках и клетках эпителия кишечника. IL-17 опосредует связь между T-клетками и гемопоэтической системой. ЦК Т-клеточного происхождения индуцирует фибробласты, которые, в результате, продуцируют IL-6, -8, ICAM-1 и G-CSF, предположительно по NF-κB-опосредованному механизму. IL-6, в свою очередь, способствует развитию колоний гранулоцитов/макрофагов, а G-CSF направляет развитие нейтрофилов. IL-17 усиливает пролиферацию частично активированных T-клеток и повышает продукцию оксида азота (NO) в хряще при остеоартрите. IL-17 может приводить к усилению АТ-зависимой гибели опухолевых клеток.
ИНТЕРЛЕЙКИН 18 (IL-18)
IL-18 – негликозилированный полипептид, у которого нет классической сигнальной последовательности. Он синтезируется в виде неактивного пропептида с м.м. 24 кДа. После протеолитического расщепления под воздействием ICE (интерлейкин-1β преобразующего энзима) или другой каспазы образуется зрелый активный пептид с м.м. 18 кДа. IL-18, также известный как IFN-γ-индуцирующий фактор (IGIF), первично был охарактеризован как потенциальный индуктор синтеза IFN-γ Т и NK клетками. Независимо от IL-12, IL-18, влияя на секрецию IFN-γ, быстро активирует клетки моноцитарно/макрофагальной системы, что ведет к активации множества антибактериальных, антиопухолевых и антивирусных ответных реакций. Сам IL-18 индуцируется стрессовыми сигналами (нейрогенными или бактериального происхождения). Считается, что индуцированное стрессом высвобождение IL-18 может вести к усилению цикла IFN-γ/IL-18: вслед за первой волной образования IFN-γ лимфоцитами, индуцированного IL-18, вновь синтезированный IFN-γ, в свою очередь, стимулирует моноциты/макрофаги, что ведет к увеличению их ICE-активности которая, в частности, приводит к образованию IL-18. IL-18 не только стимулирует синтез IFN-γ, но и модулирует его функциональную активность. Показано, что экспрессия Fas-лиганда Th1 и NK-клетками также происходит под влиянием IL-18. С другой стороны, показано, что IFN-γ участвует в активации экспрессии самого Fas. Таким образом, можно сделать вывод, что IL-18 самостоятельно (FasL) или посредством IFN-γ (Fas) стимулирует инициализацию процессов апоптоза.
ИНТЕРЛЕЙКИН 20 (IL-20)
IL-20 принадлежит семейству ЦК IL-10. Его аминокислотная последовательность приблизительно на 28% идентична IL-10 и на 76% идентична IL-20 мышей. Экспрессия IL-20 может повышаться под воздействием ЛПС. Показано, что этот ЦК инициирует каскады трансдукции с участием STAT3 и стимулирует индукцию провоспалительных генов, включая TNF-α и MCP-1. Первоначально функциональные исследования на трансгенных мышах показали, что IL-20 способен регулировать развитие кожи. Сверхэкспрессия IL-20 как у людей, так и у мышей, приводит к гиперпролиферации кератиноцитов, абнормальной эпидермальной дифференцировке и неонатальной гибели. У людей уровни IL-20 и его рецепторов повышены при псориазе. Возможно, IL-20 участвует в гемопоэзе. Он усиливает пролиферацию мультипотентных клеток-предшественников in vitro. На модельных системах показано, что IL-20 подавляет COX-2 и PGE2 и является ингибитором ангиогенеза.
ИНТЕРЛЕЙКИН 21 (IL-21)
Интерлейкин-21 - это цитокин, обладающий мощным регуляторным действием на клетки иммунной системы, включая естественные киллерные (NK) клетки и цитотоксические Т-клетки, которые могут разрушать вирусно инфицированные или раковые клетки. этот цитокин индуцирует клеточное деление / пролиферацию в своих клетках-мишенях.
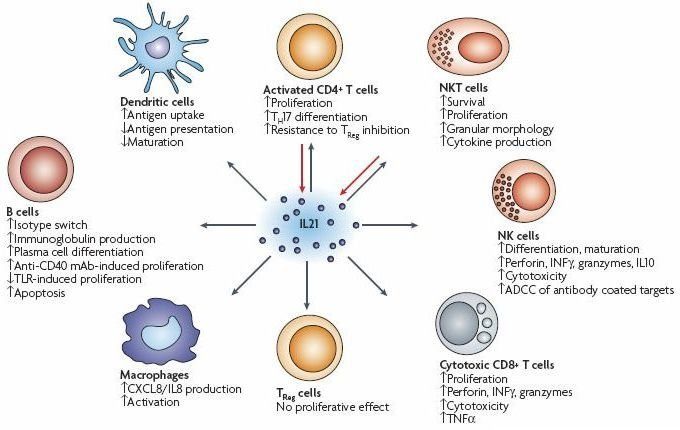
Рис. 8. Секреция IL-21 и его функции. Интерлейкин 21 (IL-21) секретируется активированными CD4+ Т-клетками, Т-фолликулярными хелперными клетками и Т-клетками-природными киллерами и способен модулировать активность большинства подгрупп лимфоцитов. Перечисленные эффекты на CD4+ T-клетки и CD8+ T-клетки наблюдались после стимуляции IL-21 вместе со стимуляцией T-клеточного рецептора или другими активирующими цитокинами (IL-2, IL-15), тогда как эффекты на NK-клетки требуют других активирующих цитокинов или активации через Fc-рецепторы. Сообщалось, что IL-21 не оказывает прямого воздействия на регуляторные T (TReg) клетки у мышей, но подавляет FOXP3 в CD4+ T-клетках человека. Другие стимуляторы В-клеток (цитокины, иммуноглобулины, агонисты толл-подобных рецепторов (TLR), лигирование CD40)) также необходимы для перечисленных эффектов на В-клетки, дендритные клетки и макрофаги. На рисунке: ADCC, антителозависимая клеточная цитотоксичность; IFN, интерферон-; mAb, моноклональное антитело; TH17, Т-хелпер 17; TNF, фактор некроза опухолей.
ИНТЕРЛЕЙКИН 22 (IL-22)
IL-22 (семейство IL-10) также известен как IL-10-подобный T-клеточный индуцибельный фактор (IL-TIF). Рецептор IL-22 экспрессируется преимущественно в поджелудочной железе, и, в меньшей степени, других тканях ЖКТ, почек и кожи. Описан растворимый рецептор – IL-22-связывающий белок (IL-22BP), который может действовать как эндогенный ингибитор активности ЦК. Показано, что IL-22 активирует сигнальные пути Jak/STAT и MAPK и усиливает продукцию белков острой фазы. IL-22 продуцируется в основном активированными Th1 и NK-клетками. Экспрессия мышиного IL-22 индуцируется в различных органах при инъекции ЛПС, что свидетельствует о возможном его участии в воспалительных ответах. Это подтверждается данными у пациентов с РА о том, что IL-22 продуцируется синовиальными фибробластами и макрофагами и может индуцировать провоспалительный ответ в синовиальных тканях. Кроме того, он стимулирует продукцию провоспалительных цитокинов и дефензинов в кератиноцитах человека.
ИНТЕРЛЕЙКИН 23 (IL-23)
IL-23 – это недавно открытый ЦК, который участвует в созревании Т-клеток памяти. Как и близкие к нему цитокины IL-12 и IL-27, IL-23 это гетеродимерный белок, субъединицы которого (p19 и p40) с м.м. ~70 кДа связаны дисульфидными связями. р19 обладает гомологией с IL-6, G-CSF и субъединицей p35 IL-12. p40, общая для IL-23 и IL-12, также обладает гомологией с рецепторами IL-6 и цилиарного нейротрофического фактора. p19 продуцируется активированными дендритными клетками, макрофагами и эндотелиальными клетками, уровень ее экспрессии повышается в ответ на бактериальные продукты. Изучение p19 показало роль IL-23 в генерации гуморального и клеточного иммунного ответа, зависимого от Т-клеток памяти. p40 продуцируется активированными дендритными клетками, макрофагами/моноцитами, микроглией, клетками костного мозга, культивируемыми в присутствии фактора стволовых клеток (c-kit лиганд) и кератиноцитами. p40 находится под контролем многих факторов транскрипции, таких как IRF-1, c-Rel, Ets-2 и Ets-подобных компонентов. При взаимодействии с перитонеальными макрофагами IL-23 стимулирует продукцию IL-1 и TNF, играя роль аутокринного воспалительного ЦК.
В настоящее время изучается роль IL-23 в патогенезе АИЗ. Показано, что IL-23 способствует образованию популяции патогенных CD4+ T-клеток, продуцирующих IL-6, -17 и TNF, а также других провоспалительных ЦК. При экспериментальной аутоиммунной энцефалопатии у мышей введение нейтрализующих АТ к IL-17 снимало аутоиммунные повреждения, опосредованные популяцией Т-клеток, развившихся под действием IL-23. Это подтверждает важность ЦК-каскада с участием IL-23 и IL-17 при АИЗ.
ИНТЕРЛЕЙКИН-31 (IL-31)
IL-31, синтезируемый активированными Th2, входит в семейство IL-6. Одна из биологических функций IL-31 – участие в патогенезе аллергических и неаллергических дерматитов, а также псориаза. Значение IL-31 состоит, прежде всего, в том, что его рецептор постоянно экспрессируется эпителиальными клетками, а также нормальными кератиноцитами эпидермиса кожи человека. Под влиянием IL-31 в кератиноцитах происходит экспрессия генов хемокинов различных семейств, в связи с чем этот цитокин рассматривается как хемоаттрактант нейтрофилов, базофилов, Т-лимфоцитов, моноцитов и др. У трансгенных мышей с резко повышенной экспрессией IL-31 наблюдалось развитие воспалительного процесса в коже, нарушения дифференцировки кератиноцитов, гиперкератоз. К этому следует добавить, что у этих трансгенных мышей при патологических изменениях кожи (выпадение шерсти, зуд и др.) наблюдалось снижение уровня IgE и IgG1 в сыворотке, что предполагает способность IL-31 включаться в патогенез неатопических дерматитов. Имеются доказательства причастности IL-31 и к тем проявлениям высыпаний на коже и экземы, в патогенезе которых важную роль играют Т-лимфоциты.
ИНТЕРЛЕЙКИН-33 (IL-33)
IL-33 – еще один член семейства IL-1, открытый путем анализа генетической базы данных по принципу родства с цитокинами этой группы. IL-33 вначале синтезируется в виде предшественника, состоящего из 270 аминокислотных остатков с м.м. около 30 кДа. Под действием каспазы-1 происходит расщепление предшественника с образованием биологически активной зрелой формы с м.м. 18 кДа. IL-33 синтезируется различными типами клеток во многих тканях. Оказалось, что рецептором для IL-33 служит известный уже достаточно давно ST2, экспрессируемый преимущественно Th2 и тучными клетками. IL-33 инициирует NF-кB и МАР-киназы, стимулирует продукцию IL-4, IL-5 и IL-13. Несмотря на структурное сходство самого IL-33 и его рецепторов с соответствующими белками семейства IL-1, этот цитокин не обладает провоспалительной активностью, но избирательно стимулирует гуморальный иммунитет, усиливая синтез Th2-цитокинов и увеличивая содержание иммуноглобулинов в плазме крови.
ОСОБЫЕ ГРУППЫ ЦИТОКИНОВ






