

Состав сооружений: решетки и песколовки: Решетки – это первое устройство в схеме очистных сооружений. Они представляют...

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Состав сооружений: решетки и песколовки: Решетки – это первое устройство в схеме очистных сооружений. Они представляют...
История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Топ:
Техника безопасности при работе на пароконвектомате: К обслуживанию пароконвектомата допускаются лица, прошедшие технический минимум по эксплуатации оборудования...
Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов...
Методика измерений сопротивления растеканию тока анодного заземления: Анодный заземлитель (анод) – проводник, погруженный в электролитическую среду (грунт, раствор электролита) и подключенный к положительному...
Интересное:
Мероприятия для защиты от морозного пучения грунтов: Инженерная защита от морозного (криогенного) пучения грунтов необходима для легких малоэтажных зданий и других сооружений...
Аура как энергетическое поле: многослойную ауру человека можно представить себе подобным...
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
В 1917 году великий шотландский зоолог Дарси Томпсон (D’Arcy Thompson) опубликовал книгу «О росте и форме», в последней главе которой представил миру свой ставший знаменитым метод трансформаций[149]. Суть метода проста: он рисовал животное на миллиметровке, а затем, изменяя масштаб согласно некоторым математическим формулам, демонстрировал превращение животного в родственное этому последнему. Миллиметровку с переменным масштабом можно представить себе листом резины. Нарисуем на ней животное, а потом растянем (или сожмем) ее. Томпсон, например, взял шесть видов крабов и нарисовал на миллиметровке один из них, Geryon. Затем «растянул» миллиметровку пятью способами и получил другие пять видов. Математические подробности нас сейчас не занимают, хотя они очень увлекательны. Но следует понять, что не так‑то трудно сделать из одного краба другого. Сам Томпсон не слишком интересовался эволюцией, но нам‑то легко представить, как должны были действовать генетические мутации, чтобы добиться таких результатов. Конечно, мы не будем считать Geryon или какой‑то из пяти других видов предком остальных. Никто из них не был таковым, и дело не в этом. Суть достижения Томпсона – в демонстрации того, что как бы ни выглядел общий предок этих крабов, трансформация такого рода может превратить любого из них (как и общего предка) в любой другой.

Ракообразные. Рисунок немецкого зоолога Э. Геккеля
Эволюция никогда не происходила посредством превращения одной взрослой особи в другую. Вспомним, что любая взрослая особь начинает существование с эмбриона. Мутация проявляется в развитии эмбриона – как изменение скорости роста частей относительно друг друга. В главе 7 мы именно так интерпретировали эволюцию черепа человека. Следовательно, если мы нарисуем человеческий череп на «резиновой миллиметровке», то преобразованиями а‑ля Томпсон сможем превратить его в череп близкого родственника, например шимпанзе, или (с большими искажениями) в череп павиана. Именно это продемонстрировал Томпсон. Заметьте, что начинать с человеческого черепа необязательно: он мог взять череп шимпанзе и получить череп человека. А еще лучше для книги по эволюции (каковой книга Томпсона не являлась) расположить на «резиновой миллиметровке» череп австралопитека и растянуть ее до черепа современного человека. Получилось бы ничуть не хуже, чем на приведенных рисунках, а с эволюционной точки зрения было бы куда осмысленней.
|
|
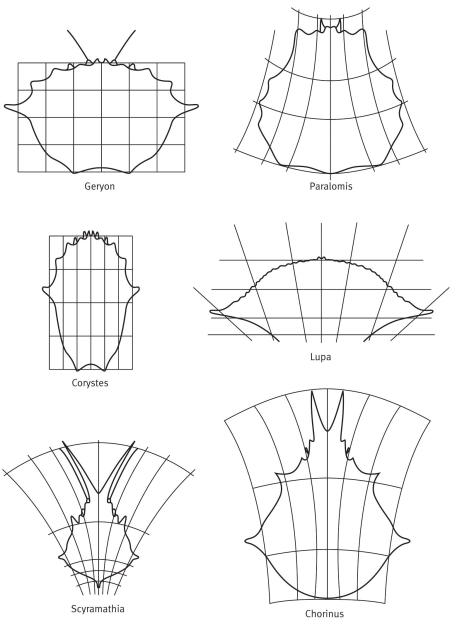
Трансформации панциря краба по Д. Томпсону
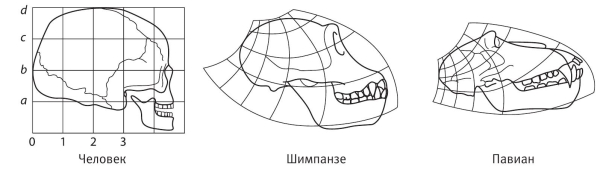
Трансформации черепа по Д. Томпсону
В начале этой главы мы познакомились с идеей гомологии на примере крыльев летучей мыши и человеческих рук. С некоторым насилием над языком я сформулировал это как совпадение скелетов при различии костей. Трансформации Томпсона позволяют сформулировать это точнее. Два органа – например, рука человека и крыло летучей мыши – гомологичны, если можно преобразованиями Томпсона получить из первого второй. Математики называют это отображение гомеоморфизмом[150].
Зоологи знали о гомологии еще до Дарвина и говорили, что, например, крыло мыши гомологично руке человека. Знай они математику лучше, они бы говорили: «гомеоморфно». После Дарвина, когда наличие у летучей мыши и человека общего предка перестало подвергаться сомнению, зоологи стали определять гомологичность в эволюционных терминах, считая гомологичными органы, унаследованные от общего предка. Термин «аналогичный» стали применять в отношении органов со сходными функциями, но без общего происхождения. Например, крыло мыши аналогично крылу насекомого и гомологично человеческой руке. Итак, если мы хотим применять гомологии как аргумент в пользу эволюционной теории, мы должны отказаться от эволюционного определения гомологии, чтобы не угодить в замкнутый круг. Поэтому нам будет пока удобнее пользоваться доэволюционным определением. Крыло мыши и рука человека гомеоморфны, поскольку одно можно получить из другого преобразованием Томпсона. Превратить таким способом крыло мыши в крыло насекомого не получится из‑за отсутствия в последнем соответствующих деталей. Широко распространенные гомеоморфизмы, которые определены без помощи эволюции, пригодны для доказательства эволюции. Нетрудно показать, как эволюция могла превратить руку одного позвоночного в руку любого другого, изменяя скорость роста частей руки у эмбриона.
|
|
С того момента, когда в 1960‑х, будучи студентом‑старшекурсником, я познакомился с компьютерами, я задавался вопросом, что сделал бы Томпсон, если бы у него был компьютер. Вопрос стал актуален в 1980‑х, с появлением доступных компьютеров с графическим дисплеем. Рисование на растяжимой подложке просто просилось на экран компьютера. Я предложил Оксфордскому университету подать заявку на грант, чтобы нанять программиста для реализации алгоритма Томпсона с удобным общедоступным интерфейсом. Мы получили деньги, и я нанял Уилла Аткинса, первоклассного программиста и биолога, который стал моим другом и советчиком. Когда он справился с широким репертуаром дисторсий Томпсона, ему уже не составляло труда научить этому программу искусственного отбора биоморфов, о которой я рассказывал в главе 2. «Игроку» предлагаются организмы, из которых он может выбрать один для «разведения». Как и прежде, процессом управляют гены, переходящие из поколения в поколение, однако теперь они управляют формой организма (растягивая или сжимая «резиновую миллиметровку», на которой он нарисован). Теоретически можно, начав с черепа австралопитека, добиться постепенного увеличения лобных долей и уменьшения длины морды по мере приближения к современному человеку. На деле же это оказалось довольно трудной задачей.
Одна из причин, на мой взгляд, по‑прежнему заключается в том, что преобразование Томпсона преображает одну взрослую форму в другую. В главе 8 я упоминал, что генетически обусловленная эволюция работает иначе. У каждого животного есть история развития. Путь от эмбриона до взрослого организма лежит через непропорциональную скорость роста отдельных частей тела. Эволюция – это не управляемое генами преобразование одного взрослого организма в другой, а управляемое генами изменение программы развития. Сэр Джулиан Хаксли (правнук Томаса и брат Олдоса[151]) осознал это, когда, вскоре после выхода первого издания книги Томпсона, попытался приспособить его метод для изучения превращения эмбриона на разных стадиях развития. Здесь мы на время, до последней главы книги, оставим метод Томпсона.
|
|
Сравнительная анатомия, как я отмечал, свидетельствует в пользу эволюционной теории даже убедительнее ископаемых остатков. Этой точки зрения придерживался и Дарвин, высказавший ее в конце главы «Взаимное родство организмов» «Происхождения видов»:
Наконец, различные группы фактов, рассмотренные в этой главе, по‑моему, столь ясно указывают, что бесчисленные виды, роды и семейства, населяющие земной шар, произошли каждый в пределах своего класса или группы от общих предков и затем модифицированы в процессе наследования, что я без колебаний принял бы этот взгляд, если бы даже он не был подкреплен другими фактами или аргументами[152].
Молекулярное сопоставление
Дарвин, конечно, не мог знать, что сравнительные доказательства станут еще более убедительными, если дополнить доступные ему анатомические сравнения молекулярной генетикой.
Так же, как скелет позвоночных остается неизменным при изменении отдельных костей, а экзоскелет всех ракообразных одинаков независимо от формы составляющих его «трубок», код ДНК одинаков у всех живых существ – меняются только отдельные гены. Это поразительный факт, яснее всех других демонстрирующий, что все живые существа происходят от общего предка. Не только генетический код, но, как мы видели в главе 8, вся генно‑белковая система, поддерживающая жизнь, одинакова у всех животных, растений, грибов, бактерий, архей и вирусов. Меняется лишь закодированная информация, сам код остается неизменным. И если посмотреть на то, что закодировано, на последовательности генов разных существ, мы снова увидим генеалогическое древо, которое получалось при сравнении скелета позвоночных, скелетов ракообразных, да даже необязательно скелета – годятся все морфологические признаки.
|
|
Допустим, мы захотели узнать, насколько близки друг другу два вида, например, ежи и обезьяны. Идеальное решение проблемы – взять полные генетические коды этих видов и сравнить их построчно и побуквенно, как библеист сравнивал бы два рукописных варианта Книги пророка Исайи. Но это дорого и отнимет много времени. Работа над проектом «Геном человека» шла десять лет, сотни человеко‑лет труда. Хотя сейчас это можно сделать намного быстрее, проект «Геном ежа» все равно будет недешевым и не слишком быстро осуществимым.
«Геном человека» – один из проектов, которые, подобно высадке на Луне или постройке Большого адронного коллайдера, вызывают у меня чувство гордости за человечество. Я рад и тому, что недавно закончены секвенирование генома шимпанзе и аналогичные проекты для других видов. Если мы и дальше будем двигаться вперед такими темпами (см. обсуждение закона Ходжкина ниже), довольно скоро сравнительный анализ и оценка родства на основе полной генетической информации станут экономически доступными. Пока же наши возможности ограничены сравнительным анализом частей генома, но и этот метод дает отличные результаты.
Для сравнения достаточно выбрать у двух видов несколько генов (или белков, строение которых известно непосредственно из генетических последовательностей) и сравнить их. Мы вскоре к этому вернемся. Однако давно известны и другие, более грубые, методы автоматизированного сравнения, которые на удивление хорошо работают. Один из них основан на использовании иммунной системы кроликов (подойдет любое другое животное, но кролики отлично справляются). Защищаясь от патогенов, иммунная система кроликов вырабатывает антитела к любому чужеродному белку[153], обнаруженному ею в крови. Как анализ антител в моей крови свидетельствует о том, что недавно меня мучил кашель, точно так же иммунный ответ кролика говорит о том, что происходило с ним в прошлом. Антитела в крови кролика – своего рода история случавшихся с ним неприятностей, включая искусственное введение белков. Если, например, впрыснуть в кровь кролика белок от шимпанзе, иммунная система выработает антитела для атаки этого белка на случай, если он снова появится в крови. Но предположим, что в следующий раз мы введем близкий, но не идентичный белок, например от гориллы. Опыт встречи с белками шимпанзе частично защитит кролика от белков гориллы, но реакция будет менее выраженной. Поможет он и от белка кенгуру, но защитная реакция будет еще слабее, поскольку кенгуру значительно дальше от шимпанзе, чем горилла. Следовательно, сила иммунного ответа может служить мерой родства нового белка с исходным, сформировавшим ответ. Именно так Винсент Сэрич и Алан Уилсон из Калифорнийского университета в Беркли показали в 1960‑х, что человек и шимпанзе гораздо ближе, чем считалось.
|
|
Есть и методы, использующие для сравнения видов гены, а не кодируемые ими белки. Один из самых старых и эффективных прямых методов – так называемая гибридизация ДНК. Именно этот метод стоит за широко известными утверждениями типа «у человека и шимпанзе более 98 % общих генов». Кстати, конкретный смысл таких цифр нуждается в уточнении: 98 % чего именно у нас общего с шимпанзе? Точный ответ зависит от размера единиц счета. Попробую пояснить это с помощью простой аналогии. Она интересна тем, что отличие от нее действительности говорит не меньше, чем сходство.
Допустим, у нас есть два варианта одной книги, и мы хотим сравнить их. Например, это каноническая Книга пророка Даниила и свиток, недавно обнаруженный в какой‑нибудь пещере на берегу Мертвого моря. Каков процент одинаковых глав в двух текстах? Скорее всего, нулевой: отличия одной буквы достаточно, чтобы различались главы. А каков процент совпадающих предложений? Вероятно, заметно выше. Процент совпадающих слов еще выше, поскольку в слове букв меньше, чем в предложении. Однако слова, отличающиеся одной буквой – все‑таки разные слова. Если мы сравним два текста, буква за буквой, то процент совпадений будет еще выше, чем со словами. Итак, оценка ничего не значит, пока мы не определим, что именно мы сравниваем: главы, параграфы, предложения, слова или буквы. Точно так дело обстоит со сравнением ДНК двух видов: если сравнивать целые хромосомы, совпадений не будет, поскольку любого отличия достаточно, чтобы хромосомы отличались.
Часто упоминаемые 98 % общего генетического материала людей и шимпанзе – это не хромосомы или целые гены, а совпадающие «буквы ДНК» (пары оснований). Однако все не так просто. Если сравнивать буквы в текстах по порядку, то вставленная или пропущенная, а не просто несовпадающая буква сдвигает все стоящие после нее буквы. Ясно, что такого рода несоответствие нас не интересует и должно быть предусмотрено. Глаз исследователя, сравнивающего варианты Книги пророка Даниила, исключает такие случаи автоматически (кстати, алгоритм такого исключения нетривиален). Как добиться того же в отношении ДНК? Настал момент оставить нашу аналогию и вернуться к ДНК, поскольку это тот случай, когда аналогия оказывается сложнее действительности.
Если ДНК постепенно нагревать, в некоторый момент (около 85 °C) связи между двумя цепочками двойной спирали разорвутся и цепочки разойдутся. Данную температуру можно считать своего рода точкой плавления. Если теперь дать цепочкам ДНК остыть, они (или их фрагменты) начнут спонтанно образовывать двойные спирали по обычным правилам связывания пар оснований. Можно было бы предположить, что повторное связывание будет происходить между теми же, полностью совместимыми, фрагментами. Так и вправду могло быть, но в действительности фрагменты ДНК найдут себе не обязательно те же пары, что прежде. Более того, если добавить фрагменты ДНК другого вида, фрагменты цепочек разных видов могут объединяться точно так же, как цепочки спирали одного вида. А почему бы и нет? Это удивительное, но очевидное следствие революционного постулата молекулярной биологии Уотсона‑Крика: ДНК – это просто ДНК. Ей нет дела, чья она: человека, шимпанзе или яблока. Фрагменты с удовольствием объединятся с любым комплементарным фрагментом, который найдут. Тем не менее сила связи непостоянна. Одиночные фрагменты цепочки ДНК связываются с другими фрагментами тем сильнее, чем более они подходят друг другу, и наоборот. Причина в том, что чем менее подходят друг другу «буквы» фрагментов (основания Уотсона‑Крика), тем больше несоответствий, тем слабее связь, как у застежки‑«молнии» с отсутствующими зубцами. Как измерить силу связи после того, как фрагменты цепочек разных видов найдут подходящие пары и свяжутся? Мы нагреем их до точки плавления! Точка плавления обычной двойной спирали с полностью соответствующими цепочками, например, ДНК человека – около 85 °C. Но когда связь слабее, например, когда цепочка ДНК человека связана с цепочкой ДНК шимпанзе, для разделения оказывается достаточно меньшей температуры. А для цепочек видов, дальше отстоящих друг от друга, например человека и рыбы либо жабы, «температура плавления» еще ниже. Разность «температур плавления» спирали, составленной из цепочек одного вида и смеси с цепочками другого, будет нашей мерой генетической близости этих видов. Правило звучит примерно так: снижение температуры на 1 °C эквивалентно 1 % несовпадающих «букв» ДНК (или отсутствию 1 % зубцов «молнии»).
Конечно, у этого метода есть сложности с нетривиальными решениями, в которые я не вдавался. Например, если смешать ДНК человека и шимпанзе, большинство фрагментов при остывании объединится с фрагментами цепочек своего вида. Как отделить «гибридную» ДНК, «температуру плавления» которой надо измерить, от двух чистых ДНК? Ответ: используя радиоактивные метки (подробный рассказ увел бы нас далеко от темы). Сейчас нам важно только то, что именно гибридизация ДНК позволяет получить оценки генетической близости наподобие 98 % для человека и шимпанзе, которые в полном соответствии с теорией уменьшаются по мере отдаления видов.
Самый современный (и затратный) метод определения генетической близости пар соответствующих генов – прямое считывание и сравнение генов тем же способом, что в проекте «Геном человека». Сравнение геномов целиком по‑прежнему стоит очень дорого, но сравнение отдельных наборов генов дает хорошие результаты и все чаще применяется на практике.
Каким бы методом измерения степени близости видов мы ни пользовались, будь то кроличьи антитела, «точки плавления» или прямое сравнение, следующий шаг будет одним и тем же. Получив числа, означающие степень близости видов, мы поместим эти числа в таблицу. Строки и столбцы – названия видов, написанные в одинаковом порядке; на их пересечении, в клетках – степень сходства. Таблица получится треугольной (рассеченный наискосок квадрат), потому что, например, сходство между человеком и собакой точно такое, как между собакой и человеком. И, если заполнить все клетки таблицы, ее половины по обе стороны диагонали окажутся зеркальным отражением друг друга.
Какой результат мы ожидаем увидеть? Эволюционная модель предсказывает, что в клетке «человек/шимпанзе» сходство будет выше, чем в клетке «человек/собака». В клетках «человек/собака» и «шимпанзе/собака» значения должны быть одинаковыми, поскольку степень родства человека с собакой та же, что у шимпанзе с собакой. То же значение должно оказаться в клетках «обезьяна/собака» и «лемур/собака». Причина в том, что человек, шимпанзе, обезьяны и лемуры связаны с собакой через одного общего предка – раннего примата, который, скорее всего, был немного похож на лемура. Так же должны соотноситься клетки «человек/кошка», «шимпанзе/кошка», «обезьяна/кошка» и «лемур/кошка», потому что кошек и собак с приматами связывает общий предок всех хищников. Во всех клетках, связывающих, например, кальмара с млекопитающими, должно стоять примерно одно и то же число, причем маленькое. И неважно, о каком млекопитающем идет речь: они одинаково далеки от кальмара.
Но это теория, а практика иногда сильно от нее отличается. Если мы увидим противоречие, оно будет аргументом против эволюционной теории. Однако в действительности (с учетом статистической погрешности) мы наблюдаем именно то, что предсказывает эволюционная теория. Иными словами, если приписать значения сходства между парами видов к ветвям нашего дерева, то все сойдется. Конечно, абсолютного совпадения нет, но численные оценки в биологии редко даются с точностью выше некоторого приближения.
Доводы сравнительного анализа ДНК или белков позволяют сделать вывод о том, какие пары видов являются близкородственными. Однако сила этого метода как доказательства эволюционной теории заключается в возможности построить дерево генетического родства по каждому гену. Важнейший результат заключается в том, что все такие деревья во многом одинаковы. Именно такой результат и должен был получиться, если перед нами генеалогическое древо. И совсем не то, что мы ожидали, если бы Творец выбирал подходящие белки то там, то здесь, вне зависимости от первоначального носителя белка.
Первое масштабное исследование этого рода было выполнено группой новозеландских генетиков под руководством профессора Дэвида Пенни[154]. Группа Пенни выбрала пять генов, которые пусть не идентичны у всех млекопитающих, но достаточно схожи для того, чтобы у всех называться одинаково. Это гены гемоглобина А, гемоглобина В (он придает крови красный цвет), фибринопептида А, фибринопептида В (фибринопептиды отвечают за свертывание крови) и цитохрома С, играющего важную роль в биохимии клетки. Для сравнения были выбраны одиннадцать млекопитающих: макак‑резус, овца, кенгуру, крыса, кролик, собака, свинья, человек, лошадь, корова и шимпанзе.
Пенни с коллегами подошли к задаче статистически. Они решили оценить вероятность того, что две молекулы могут дать одно и то же генеалогическое древо случайно. Поэтому они перебрали все деревья, которые можно построить для выбранных одиннадцати видов. Это число оказалось на удивление большим. Даже если ограничиться бинарными деревьями (в каждом узле ствол делится на две ветви, то есть дает двух потомков), то количество вариантов превысит 34 миллиона. Ученые терпеливо перебрали эти деревья, сравнив каждое с остальными 33999999 вариантами. То есть, конечно, они не делали этого: ушло бы слишком много времени. Вместо этого они придумали изощренный статистический эквивалент этих чудовищных по объему вычислений.
Метод приблизительно такой. Возьмем первые пять генов, например, гемоглобина А (я буду использовать здесь название белка, который кодируют гены) и найдем среди миллионов деревьев самое экономное для гемоглобина А. Под экономным понимается дерево, которое требует минимального числа эволюционных изменений. Например, все деревья, где кенгуру ближе к человеку, чем к шимпанзе, далеки от экономного: они требуют очень большого числа изменений, чтобы у кенгуру и человека нашелся близкий общий предок. Если бы гемоглобин А умел говорить, он рассказал бы примерно следующее:
Это очень неэкономное дерево. Оно требует не только огромного числа мутаций, чтобы получить два таких разных, но в то же время близкородственных организма, как кенгуру и человек, но еще и огромного числа мутаций, чтобы человек и шимпанзе оказались столь похожими с моей, гемоглобиновой, точки зрения. Я против этого варианта.
В отношении каждого из 34 миллионов деревьев гемоглобин А вынес свой вердикт, отобрав в итоге десятка полтора самых экономных деревьев. О каждом из них он отозвался бы приблизительно так:
В этом варианте человек и шимпанзе – близкие родственники, коровы и овцы также, а кенгуру находятся где‑то на своей ветке. Это хорошее дерево, потому что мне не придется выполнять почти никакой мутационной работы для того, чтобы объяснить эволюционные изменения. Это очень экономное дерево. Я голосую «за».
Конечно, было бы приятно, если бы гемоглобин А и остальные гены «проголосовали» за одно и то же экономное дерево, но это было бы слишком. Понятно, что среди 34 миллионов деревьев найдется несколько достаточно симпатичных гемоглобину А.
Однако не будем забывать о гемоглобине В, цитохроме С и других белках. Когда все они голосовали за своих кандидатов из 34 миллионов деревьев, можно было бы ожидать, что их вкусы окажутся разными. Могло, например, оказаться, что с точки зрения цитохрома С человек и вправду гораздо ближе к кенгуру, чем к шимпанзе. В отличие от гемоглобина А, цитохром С мог бы, например, сказать, что ему не составит больших (мутационных) затруднений превратить овцу в обезьяну или сделать коров и кроликов близкими родственниками. Если придерживаться креационистской гипотезы, этому ничто не препятствует. Но Пенни и его коллеги обнаружили, что все пять генов голосуют практически одинаково (более того, они оценили вероятность того, что результат мог случайно оказаться таким же). Все гены «проголосовали» за одни и те же деревья, чего следовало ожидать эволюционисту, предполагающему существование одного дерева, связывающего одиннадцать животных в генеалогическое древо – древо эволюционных изменений. Самое любопытное, что дерево, победившее в «голосовании» генов, оказалось именно тем вариантом, который зоологи получили независимо, на основании анатомических и палеонтологических данных.
Результаты исследования Пенни были опубликованы в 1982 году, то есть довольно давно. С тех пор было проделано огромное число детальных исследований на основе конкретных генетических последовательностей разных животных и растений. Консенсусное древо, которое удовлетворяет всех участников «голосования», теперь охватывает куда больше пяти генов и одиннадцати видов оригинального исследования Пенни. Это был просто изящный пример, оказавшийся очень сильным аргументом из‑за примененной статистической методологии.
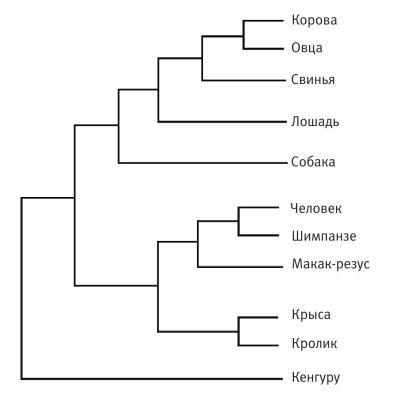
Древо эволюционных изменений по Д. Пенни
Объем генетических последовательностей, накопленный за последнее время, вообще устраняет всякие сомнения. Результаты сравнения генетических деревьев куда более убедительно, чем данные палеонтологии (сами по себе убедительные), сходятся стремительно и безусловно к единому великому древу жизни. На иллюстрации изображено дерево млекопитающих, которых изучал Пенни и его коллеги, но уже с учетом консенсусного голосования гигантского количества частей их генома. Именно постоянная согласованность результатов исследований разных генов позволяет нам быть уверенными не только в точности построения самого дерева, но и в самом факте эволюции.
Если молекулярная генетика и дальше будет развиваться экспоненциально, то к 2050 году секвенирование генома любого существа будет занимать не больше времени и стоить не дороже, чем сейчас измерение температуры или кровяного давления. Почему я считаю, что генетические технологии развиваются по экспоненте? Можно ли измерить скорость их развития? В кибернетике существует аналогичное наблюдение – закон Мура, названный в честь одного из основателей компании «Интел». Поскольку существуют разные метрики производительности компьютеров, есть несколько взаимосвязанных формулировок этого закона. Одна из них гласит: количество элементов, которые можно разместить на интегральной микросхеме, удваивается в течение 18–24 месяцев. Это эмпирический закон, выведенный на основе накопленных данных. Закон Мура выполняется уже около пятидесяти лет и, по мнению многих экспертов, останется в силе по меньшей мере еще несколько десятилетий. Другие формулировки закона Мура относятся к скорости вычислений, объему памяти и другим характеристикам в расчете на единицу стоимости, и все они носят экспоненциальный характер. Еще Дарвин продемонстрировал, что экспоненциальный рост ведет к удивительным результатам. С помощью своего сына Джорджа, математика, он показал, что слон, довольно медленно размножающееся животное, всего за несколько столетий неограниченного экспоненциального размножения может заполнить своими потомками планету. Конечно, слоны не размножаются экспоненциально: их размножение ограничено соревнованием за пищу и пространство, болезнями и другими факторами. Собственно, именно это – в какой момент естественный отбор начинает действовать – хотел показать Дарвин.
Закон Мура действует по крайней мере последние полвека. Никто не знает почему, но компьютерная мощность увеличивается экспоненциально, а дарвиновские слоны размножаются по экспоненте только в теории. Я предположил, что аналогичный закон должен действовать и в отношении генной инженерии. Я поделился своим предположением с Джонатаном Ходжкином, оксфордским профессором генетики, моим бывшим студентом. К моей радости, он уже успел об этом подумать и даже провел необходимые вычисления, оценив стоимость секвенирования ДНК стандартного размера в 1965‑м, 1975‑м, 1995‑м и 2000 году. Я представил, сколько ДНК можно секвенировать за тысячу фунтов стерлингов. Полученные цифры отложил на графике в логарифмическом масштабе, что обычно наглядно для экспоненциально растущих величин (на логарифмической шкале они выглядят как прямые линии). Как и должно быть, все четыре точки Ходжкина лежали почти на прямой. Я подобрал к ним прямую (методом линейной регрессии) и экстраполировал ее на будущее. Когда книга отправлялась в типографию, я показал это место в рукописи профессору Ходжкину и узнал о последнем известном ему достижении в этом направлении – секвенировании в 2008 году генома утконоса. Это само по себе очень интересно, поскольку утконос занимает стратегическое положение на древе жизни: наш общий предок жил 180 миллионов лет назад, почти в три раза раньше, чем вымерли динозавры. Я добавил утконоса в график (отмечен звездочкой) и порадовался, что он оказался близко к предполагаемому положению.
Наклон прямой, построенной на основании зависимости, которую я назвал законом Ходжкина (без его разрешения), чуть более пологий, чем у закона Мура, а время удвоения – чуть больше двух лет против чуть менее двух лет для закона Мура. Генная инженерия сильно зависит от компьютерных технологий, так что естественно ожидать сходства обоих законов.
Стрелками показаны размеры генома разных существ. Проведя прямую от стрелки до пересечения с прямой Ходжкина, можно узнать, когда этот геном можно будет секвенировать за тысячу нынешних фунтов стерлингов. Генома дрожжей нужно ждать до 2020 года. В случае генома млекопитающего (с этой точностью все млекопитающие одинаково сложны) моя оценка – чуть раньше 2040 года. Это фантастическая перспектива: всеобъемлющая база данных ДНК всех животных и растений всех царств за приемлемые деньги. Детальное сопоставление ДНК закроет все бреши в знаниях об эволюционном родстве видов и даст нам древо жизни всех живых существ[155]. И как мы только будем рисовать его? Никакого листа не хватит.
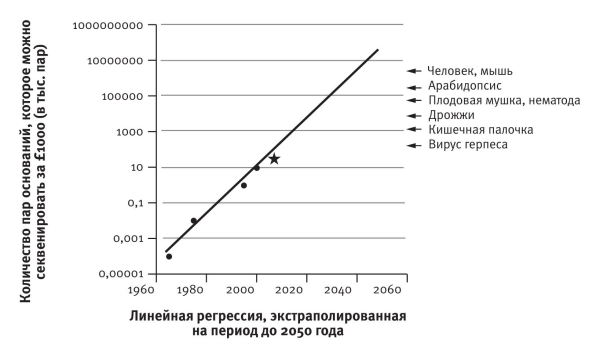
Закон Ходжкина
Самый масштабный проект в этой области был предпринят группой Дэвида Хиллиса (брата разработчика суперкомпьютеров Дэнни Хиллиса). График Хиллиса позволяет представить дерево в более компактном виде посредством наложения на круг. Разрыв, где концы встречаются, почти не виден: он находится между бактериями и архебактериями. Взглянем, например, на сокращенную версию, которую зоолог Клэр Д’Альберто из Мельбурнского университета вытатуировала у себя на спине (интерес Клэр к зоологии поверхностным точно не назовешь). На татуировке (цветная вклейка 25) небольшая выборка – всего 86 видов: именно столько изображено терминальных веточек. Глядя на разрыв на круговом графике, можно представить, что круг развернули.
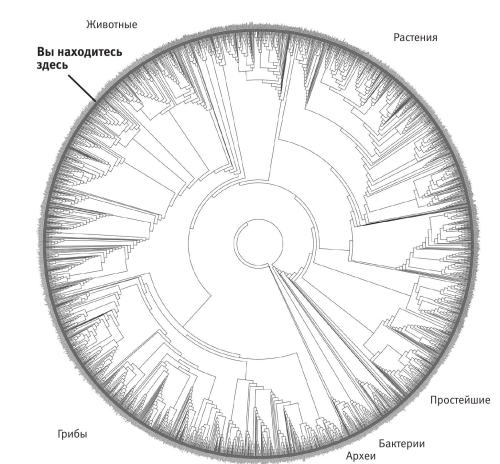
График Хиллиса
Несколько иллюстраций с края – бактерии, простейшие, растения, грибы и четыре других типа животных – выбраны для наглядности. Позвоночные представлены морским драконом – удивительной рыбой, которая защищается, маскируясь под водоросли. График Хиллиса устроен точно так же, однако включает три тысячи видов. Их названия нанесены с внешней стороны круга, но так мелко, что их трудно прочесть, поэтому вид Homo sapiens отмечен особо. Даже столь подробный чертеж покрывает лишь небольшую часть древа жизни: ближайшие родственники человека на нем – крысы и мыши. Число млекопитающих пришлось сильно сократить, чтобы остальные ветви вошли до той же глубины. Попробуйте вообразить себе дерево с десятью миллионами видов вместо трех тысяч у Хиллиса. И, замечу, десять миллионов – не самая вызывающая оценка числа существующих видов. Советую скачать древо Хиллиса на его сайте[156], напечатать и повесить на стену, если, конечно, найдете лист бумаги шириной не менее 137 см.
Молекулярные «часы»
Беседуя о молекулах, мы не должны забывать неоконченную историю из главы об эволюционных часах. Там мы обсуждали годичные кольца, разные виды радиоактивных «часов», но отложили обсуждение так называемых молекулярных «часов». Теперь время пришло.
Принцип действия молекулярных «часов» основан на том, что эволюция действительно происходит, причем идет с постоянной скоростью относительно геологического времени, калиброванного ископаемыми останками, возраст которых в свою очередь установлен радиоуглеродным анализом. Так же, как работа водных часов основана на предположении о постоянной скорости вытекания из сосуда воды, а механических – на постоянстве периода колебаний маятника, молекулярные часы предполагают, что некоторые эволюционные процессы протекают с постоянной скоростью. Для калибровки этой скорости можно использовать любые хорошо документированные свидетельства эволюции, например, ископаемые находки, датированные радиоуглеродным методом. После калибровки молекулярные «часы» можно применять для датирования других, не столь хорошо документированных частей эволюционной истории. Например, их можно использовать для животных, у которых нет твердого скелета и которые редко оставляют записи в палеонтологической летописи.
Идея заманчивая. Но что дает нам основание считать эволюционные процессы протекающими с постоянной скоростью? Множество фактов свидетельствует о высокой изменчивости ее скорости. Когда‑то Холдейн предложил измерять скорость эволюции в дарвинах. Пусть какая‑то измеряемая характеристика организма за время эволюции устойчиво изменяется: например, растет средняя длина ноги. Если за миллион лет нога выросла в е раз (2,71828… – число, выбранное исходя из математического удобства, но сейчас это не важно)[157], скорость эволюционных изменений равна одному дарвину. По оценке Холдейна, скорость эволюции лошади составляет 40 миллидарвинов, а скорость эволюции домашних животных, подвергающихся искусственному отбору, оценивается в килодарвинах. Скорость эволюции гуппи, пересаженных в водоем без хищников (см. главу 5), составляет около 45 килодарвинов. Эволюция «живых ископаемых», таких как брахиоподы Lingula, протекает со скоростью микродарвинов. Итак, скорость эволюции признаков, которые можно увидеть и измерить, таких как ноги или клювы, весьма изменчива.
Почему же мы рассчитываем использовать скорость эволюции в качестве часов? На помощь спешит молекулярная генетика. На первый взгляд не очень понятно, чем она может помочь. Когда измеряемый признак меняется (например, растущая нога), мы видим в этом внешнее проявление генетических изменений. Как же может случиться, что скорость изменений на генетическом уровне достаточно устойчива для того, чтобы служить часами, а скорость изменений их проявлений переменна? Генетические изменения, проявляющиеся в видимых изменениях, – лишь верхушка айсберга, на которую сильно влияет естественный отбор. Большая часть изменений на молекулярном уровне нейтральна к отбору и не сказывается на способности организма к выживанию, так что мы можем ожидать постоянства скорости этих изменений для каждого гена и, следовательно, их пригодности в роли часов.
Когда нейтральная теория молекулярной эволюции была предложена великим японским генетиком Кимура Мотоо и другими учеными, она была весьма противоречивой. Сейчас общепринятой является ее версия, которой я, не вдаваясь в детали, и буду придерживаться. Поскольку у меня репутация архиадаптациониста (якобы упорствующего в том, что естественный отбор – это основная, даже единственная движущая сила эволюции), можете быть уверены, что если я поддерживаю эту теорию, с ней вряд ли кто‑то еще не согласен[158].
Нейтральной считается измеримая методами молекулярной генетики мутация, которая не подвержена ни негативному, ни позитивному естественному отбору. «Псевдогены» нейтральны по вполне понятной причине: это гены, когда‑то выполнявшие определенные функции, а после ставшие невостребованными, оттесненные на периферию и больше не участвующие ни в транскрипции, ни в трансляции. С точки зрения благополучия организма они не существуют. Однако для ученого они еще как существуют: именно они нужны для эволюционных «часов». В эмбриологии «псевдогены» – не единственный нетранслируемый класс генов. Есть и другие классы, подходящие для молекулярных «часов». Для чего «псевдогены» подходят лучше всего, так это для того, чтобы смущать креационистов: даже необузданной фантазии этих людей недостаточно для объяснения того, зачем, если не для того, чтобы дурачить нас, Творец создал «псевдоген», который ничего не делает, но очень похож на престарелую версию когда‑то активного гена.
Большая часть генома (у людей – около 95 %) вроде бы ни для чего не нужна. Но нейтральная теория пр<
|
|
|

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

История развития пистолетов-пулеметов: Предпосылкой для возникновения пистолетов-пулеметов послужила давняя тенденция тяготения винтовок...

Археология об основании Рима: Новые раскопки проясняют и такой острый дискуссионный вопрос, как дата самого возникновения Рима...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!