В основе нелепых требований предъявить «недостающие звенья» лежит средневековый миф, который властвовал над умами ученых мужей до появления Дарвина и который поныне продолжает многих смущать. Я говорю о концепции Лестницы существ, в соответствии с которой все во Вселенной расположено иерархически, на ступеньках гигантской лестницы – на самом верху Бог, после архангелы, после ангелы разных чинов, после люди, после животные, растения и так далее, вплоть до камней и других объектов неживой природы. Учитывая, что этот миф относится к временам, когда расизм цвел пышным цветом, вряд ли надо уточнять, что тогда и не все люди считались достойными занимать одну и ту же ступень. Конечно, мужчины горделиво стояли ступенью выше женщин своего ранга (и поэтому в начале абзаца я написал: «над умами ученых мужей»). Но когда на сцене появилась эволюционная теория, больше всего путаницы породили устаревшие представления об иерархии внутри животного царства. Казалось логичным, что «низшие» животные эволюционировали в «высших». Следовательно, мы вправе ожидать обнаружения «связующих звеньев» между ними, отражающих восхождение по иерархической лестнице. Лестница с провалами выглядит неубедительно. И именно этот образ – лестницы с недостающими ступенями – лежит в основе скепсиса по отношению к «недостающим звеньям». Но, как я сейчас покажу, миф о Лестнице существ целиком ошибочен и антиэволюционен.
Словосочетания «низшие животные» и «высшие животные» так легко слетают с нашего языка. Так же легко, казалось бы, они должны вписываться в эволюционное мышление – и тем неожиданнее осознание того, что эти понятия глубоко антиэволюционны. Казалось бы, мы всегда понимали, что шимпанзе – существо более сложное, «высшее», в то время как дождевые черви – «низшие» существа. Мы думаем, будто мы знаем, что это значит – и эволюция, казалось бы, еще лучше проясняет смысл этих понятий. Однако это не так. На самом деле вообще не ясно, имеют ли смысл эти слова. Если же они значат что‑либо, то значений столько и они такие разные, что в итоге разделение животных на «низших» и «высших» только запутывает или даже оказывается откровенно вредным.
Перечислим несколько более или менее ошибочных смыслов, которые можно вложить во фразу: обезьяна – «высшее» в сравнении с дождевым червем животное.
1. «Обезьяны в процессе эволюции произошли от дождевых червей». Это неверно, как неверно и то, что человек произошел от шимпанзе. У обезьяны и дождевого червя, шимпанзе и человека есть общий предок.
2. «Общий предок дождевого червя и обезьяны был куда больше похож на дождевого червя, чем на обезьяну». Это куда лучше! Кроме того, слово «примитивный» можно использовать как почти точный термин, отражающий степень сходства с общим предком. Очевидно, что по этому признаку некоторые из современных животных «примитивнее» других. Тогда более «примитивным» из двух видов будет тот, который изменился меньше со времени их общего предка (поскольку, как мы помним, все виды имеют общего предка – надо только отойти достаточно далеко назад по эволюционному пути). Если же оба вида изменились примерно в одинаковой степени, слово «примитивный» использовать не стоит.
Не лишним будет сделать небольшое отступление. Сходство двух видов очень сложно измерить. И, более того, нет никаких причин предполагать, что общий предок каких‑либо двух современных животных непременно был сильнее похож на одно из них, чем на другое. Возьмем, например, сельдь и кальмара. Вполне возможно, что один из них больше похож на их общего предка, чем другой, но из этого не следует, что так и было. У обоих животных было равное количество времени на эволюционные изменения. Следовательно, с точки зрения эволюциониста, ни одно из современных животных не более примитивно, чем другое. Нам следует ожидать, что оба животных со времени общего предка изменялись с одинаковой скоростью, но в разных направлениях. Мы знаем, что это ожидание часто не соответствует действительности (например, в случае обезьяны и дождевого червя), но в принципе для каких‑то животных оно может подтвердиться. Более того, даже отдельные части тела животного могут эволюционировать с разной скоростью. Животное может быть примитивным от пояса и ниже, а выше пояса очень развитым. У одного животного может быть более примитивная нервная система, а у другого, например, скелет. Отметим, что «примитивный» («более напоминающий предков») совершенно не означает «простой» («менее сложно устроенный»). Конское копыто устроено проще, чем наша нога, поскольку имеет один палец вместо пяти, однако человеческая ступня примитивнее, потому что наш общий с лошадьми предок имел, как и мы, пять пальцев. Следовательно, лошади в процессе эволюции изменились сильнее.
3. «Обезьяны умнее (симпатичнее, или имеют более длинный геном, или более сложное строение тела и так далее), чем кольчатые черви». Подобный зоологический снобизм приводит в науке к кошмарной путанице. Можно представить себе множество «шкал» развитости животных. Однако животные, получившие высокий балл в одном случае, совершенно не обязательно окажутся настолько же развитыми в другом. У млекопитающих, например, мозг крупнее, чем у саламандры, однако у некоторых саламандр существенно крупнее геном.
4. «Обезьяны похожи на людей сильнее, чем кольчатые черви». Да, это так. И что же? Почему мы должны брать человека за меру, которой мы измеряем другие организмы? Пиявка, к примеру, могла бы с негодованием отметить, что дождевые черви обладают огромным преимуществом: они ближе к пиявке, чем к человеку! Несмотря на традиционное помещение людей между животными и ангелами на Лестнице существ, нет подтверждений распространенному убеждению, что эволюция «нацелена» на выведение человека или что человек – венец творения. В худшем случае все опускается до вечного ворчания вроде: «Если шимпанзе эволюционировали в человека, то почему мы до сих пор видим их?» Увы, это не анекдот. Сотни раз я слышал этот вопрос, причем иногда от людей, на первый взгляд хорошо образованных[75].
5. Обезьяны (и прочие «высшие» животные) приспособлены для выживания лучше, чем дождевые черви (и прочие «низшие» животные). В этом нет не только правды, но и смысла. Все существующие виды дожили до наших дней. Многие обезьяны, например, изящные львиные игрунки, находятся на грани вымирания. Они приспособлены к выживанию куда хуже дождевых червей. Крысы и тараканы, которых многие считают «низшими» животными, процветают, в то время как «высшие» гориллы и орангутаны близки к исчезновению.
Надеюсь, что я достаточно ясно высказался об абсурдности попыток представить современные виды как «низшие» и «высшие», а также об антиэволюционности такого подхода. Да, можно придумать массу иерархических моделей, и некоторые из них в отдельности иногда могут оказаться полезными. Однако они практически не коррелируют друг с другом, и, тем более, ни у одной нет права называться «шкалой эволюционного развития». Мы с вами уже были свидетелями соблазнительности вопроса, почему в мире нет лягузьян. Кроме того, трагическим наследием концепции Лестницы существ являются распространенный вопрос: «Где промежуточные звенья между большими группами животных?» и (что весьма прискорбно) склонность многих эволюционистов отвечать на подобные вопросы, поднимая на щит конкретные ископаемые, например, археоптерикса – знаменитое «промежуточное звено между рептилиями и птицами». Тем не менее, у истории археоптерикса есть важная подоплека, и на ней я хотел бы остановиться. Рассмотрим случай с археоптериксом как частное проявление ситуации.
Исторически зоологи подразделяют позвоночных животных на классы – обширные категории наподобие млекопитающих, птиц, рептилий и амфибий. Зоологи, относящиеся к кладистам[76], настаивают, что истинный класс должен состоять из животных, имеющих общего предка, принадлежащего к тому же классу и не имеющего потомков вне этой группы. Примером полноценного класса являются птицы[77]. Все птицы происходят от общего предка, которого также можно считать птицей, поскольку он обладал всеми ключевыми особенностями современных птиц: он имел перья, крылья, клюв и так далее. А вот животные, называемые рептилиями, в этом смысле истинным классом не являются. По крайней мере в классической таксономической системе класс рептилий не включает птиц, поскольку они составляют отдельный класс. Однако некоторые рептилии (например, крокодилы и динозавры) ближе к птицам, чем к другим рептилиям (ящерицам и черепахам). Более того, некоторые динозавры ближе к птицам, чем к другим динозаврам! Таким образом, рептилии – это искусственный класс, поскольку птицы из него искусственно исключены. Если бы мы захотели сделать из рептилий истинный класс в кладистическом понимании, птиц пришлось бы объединить с рептилиями. Зоологи‑кладисты вообще избегают термина «рептилии», предпочитая разделять эту группу на архозавров (крокодилы, динозавры и птицы), лепидозавров (змеи, ящерицы и редкий новозеландский род Sphenodon) и панцирных (морских и сухопутных черепах). Не‑кладисты по‑прежнему используют термин «рептилии», находя его удобным, даже если он исключает птиц.
Но что есть такое в птицах, что заставляет нас выделить их в отдельный класс, когда они, в сущности, просто одна из ветвей рептилий? Ответ прост: ближайшие родственники птиц среди рептилий давно вымерли, а птицы, единственные в своем роде, продолжили победоносное шествие сквозь время. Непосредственные родственники птиц – динозавры. Если бы выжило несколько потомственных линий динозавров, птицы ничем не выделялись бы и не были бы подняты до уровня отдельного класса млекопитающих, и мы бы не вопрошали, где связующее звено между рептилиями и птицами. Археоптерикс оставался бы прекрасным экспонатом палеонтологического музея и не играл бы нынешней «звездной» роли, исполняемой по первому требованию показать «промежуточные звенья», – требованию, как мы видели, пустому. Если бы карта вымирания видов легла по‑другому, вокруг нас бегала, летала и плавала бы масса динозавров, среди которых были бы пернатые, летающие, клювастые динозавры, именуемые птицами. И, естественно, в последнее время обнаруживается все больше окаменелых остатков пернатых динозавров, что делает очевидным отсутствие самой проблемы «недостающего звена», решением которой считался археоптерикс. Перейдем теперь к некоторым наиболее значимым эволюционным переходам, в которых, как считалось, имеются «недостающие звенья».
Из моря на сушу…
Нелегко представить себе сильнее меняющее уклад жизни событие, чем полет человека в космос и выход животных на сушу. Переход от одной среды к другой требует радикальных перемен во всех частях и системах тела. Жабры, превосходно поглощающие кислород из воды, в атмосфере бесполезны, а легкие бесполезны в воде. Способы передвижения, в воде ловкие, быстрые и маневренные, неприемлемы на суше – и наоборот. Не случайно появились пословицы «как рыба, вытащенная из воды» и «плавает как топор». И не случайно «недостающие звенья» в палеонтологической летописи, относящиеся к этому периоду, вызывают повышенный интерес.
Если нырнуть поглубже во время, мы обнаружим, что когда‑то все живые существа обитали в море, этой соленой и мокрой колыбели. Время от времени в эволюционной истории случалось, что предприимчивые особи из разных групп организмов выходили на сушу, иногда даже в самые сухие из пустынь, и уносили с собой морскую воду в виде собственной крови и клеточных жидкостей. Кроме рептилий, птиц, млекопитающих и насекомых, на суше преуспели скорпионы, улитки, ракообразные (сухопутные крабы и мокрицы), кивсяки и сороконожки, пауки со своими родичами, по меньшей мере три типа червей. Не следует забывать также об источнике углерода – растениях, выигравших авангардный бой, без победы в котором не смогла бы произойти ни одна из упомянутых миграций.
Нам невероятно повезло в том, что палеонтологическая летопись содержит записи о пути исхода рыб на сушу. Как, кстати, и обратного процесса, в течение которого предки китов и дюгоней много позднее бросали с трудом завоеванную сушу и возвращались в море. Звенья, считавшиеся недостающими, в настоящий момент обнаружены и украшают наши музеи.
Говоря, что рыбы выбрались на сушу, следует помнить прежде всего о том, что рыбы, как и рептилии, не представляют собой естественную группу. Рыбы – это все позвоночные, за исключением вышедших на сушу. Поскольку эволюция позвоночных на ранних стадиях происходила в море, не удивительно, что там и осталась большая часть ветвей этой группы. И, даже если они очень далеки от других рыб, мы называем их рыбами. Форель и тунец ближе к человеку, чем к акуле, однако и тех, и других мы считаем рыбами. Двоякодышащие и латимерии ближе к людям, чем к тунцу и форели (и, конечно, акуле), однако и их называют рыбами. Даже акулы ближе к человеку, чем к миногам и миксинам (единственным ныне живущим представителям некогда процветающей группы бесчелюстных рыб). Но и их мы называем рыбами. Позвоночные, предки которых никогда не выходили на землю, выглядят как рыбы, плавают как рыбы (в отличие, например, от дельфинов, которые плывут, изгибая позвоночник вверх и вниз, а не вправо‑влево) и, полагаю, на вкус как рыба.
Из примеров с птицами и рептилиями видно, что для эволюциониста естественная группа животных – это такая группа, все члены которой ближе друг к другу, чем к любому организму, не являющемуся членом группы. Птицы – естественная группа, поскольку их последний общий предок не имеет других потомков, кроме птиц, а рыбы и рептилии естественными группами не являются. Последний общий предок всех рыб является также общим предком многих других организмов. Если отталкиваться от акул, то мы, млекопитающие, принадлежим к группе, включающей также современных костистых рыб (в противоположность хрящевым, например акулам). Если отталкиваться от костистых, или лучеперых рыб (лосось, форель, тунец, рыбаангел, вообще все рыбы, которых вы видели, кроме акулы), то мы принадлежим к группе, к которой также относятся все наземные позвоночные, а также так называемые лопастеперые рыбы. Именно от лопастеперых рыб произошли все наземные позвоночные, и потому нам стоит обратить особое внимание на их плавники.
К нашему времени из лопастеперых сохранились только целаканты и двоякодышащие рыбы (ну и, конечно, все наземные позвоночные). Лопастеперыми их называют потому, что их мясистые плавники больше напоминают ноги, чем типичные лучевые плавники хорошо знакомых нам рыб. Недаром книга известного специалиста по целакантам Дж. Л. Б. Смита, южноафриканского ихтиолога, представившего их широкой общественности после того, как в 1938 году при тралении поймали первого живого представителя вида, называлась «Старина Четвероног». Смит писал: «Не нужно особого воображения, чтобы представить себе, какой переполох вызвало бы внезапное появление перед нами исполинского динозавра! Вообще обнаружить какой‑нибудь фрагмент далеких эпох считается целым событием. И пусть целакант ростом не с динозавра, его появление во многом еще более поразительно». Целаканты были хорошо известны в виде отпечатков и ископаемых остатков, однако считалось, что они вымерли еще во времена динозавров.
Смит с чувством описал свою первую встречу с целакантом. Взглянуть на этот потрясающий улов и высказать экспертное мнение Смита попросила Марджори Куртенэ‑Латимер, обнаружившая рыбу (которую Смит позднее назвал латимерией):
После утомительного путешествия мы прибыли в Ист‑Лондон[78] и сразу же направились в музей. Мисс Латимер куда‑то вышла, но сторож провел нас внутрь, и мы увидели… целаканта! Силы небесные! Хоть я и был подготовлен, меня в первый миг словно ошеломило взрывом, ноги подкосились, тело стало чужим. Я будто окаменел. Да, никаких сомнений: чешуя, кости, плавники – самый настоящий целакант. Точно внезапно ожила рыба, умершая 200 миллионов лет назад. Забыв обо всем на свете, я смотрел не отрываясь на чучело, потом робко подошел ближе, коснулся его, погладил… Жена молча следила за мной. Вошла мисс Латимер и тепло с нами поздоровалась. Только тут ко мне вернулся дар речи. Не помню точно, о чем я говорил, но смысл моих слов сводился к тому, что не может быть никаких сомнений – это он, да‑да, целакант, совершенно точно! Даже я не мог больше сомневаться[79].
Целаканты ближе к нам, чем к большинству рыб. Они несколько изменились со времен нашего последнего общего предка, но не слишком сильно для того, чтобы, по общему признанию и с точки зрения рыбака, быть выделенными из группы рыб. Но они, как и двоякодышащие, действительно ближе к нам, чем к форели, тунцу и вообще большинству рыб. Целаканты и двоякодышащие – примеры так называемых «живых ископаемых».
Тем не менее мы не происходим ни от двоякодышащих, ни от целакантов. С двоякодышащими нас объединяет предок, который был больше похож на рыбу, чем на человека. Но, если честно, он и на рыбу‑то не был похож. Двоякодышащие – живые ископаемые, но все же они очень далеки от наших общих предков. В поисках этих существ нам надо обратиться не к живым, а к настоящим ископаемым, в особенности из девонских слоев: там зафиксирован переход от живших в воде рыб к первым позвоночным, освоившим сушу. Мы были бы чересчур самонадеянны, если бы рассчитывали найти наших предков даже среди ископаемых, но все же вполне можно найти остатки организмов, достаточно близких к нашим предкам, и по их строению понять, на кого наши предки были похожи.
Среди пробелов в палеонтологической летописи хорошо известен так называемый пробел Ромера. Он назван в честь известного американского палеонтолога Альфреда Шервуда Ромера и представляет собой временной отрезок с конца девонского периода (примерно 360 миллионов лет назад) до первой трети карбона (около 340 миллионов лет назад). После пробела Ромера мы находим безошибочно определяемых амфибий, расплодившихся в болотах, богатое разнообразие саламандроподобных животных, некоторые из которых были размером с крокодила, напоминая его и своим обликом. Вообще, это был настоящий век гигантов: в те времена жили даже стрекозы с размахом крыльев больше моей руки – самые крупные из обитавших на планете насекомых[80]. Каменноугольный период, начавшийся примерно 340 миллионов лет назад, для амфибий представлял собой аналог эры динозавров. Но перед ним – провал, пустота, пробел Ромера. А прежде этого пробела мы вместе с Ромером видим только рыб – лопастеперых рыб, живших в воде. Где же «промежуточные звенья»? И что заставило их выйти на сушу?
В далекие времена студенчества в Оксфорде мое воображение будоражили лекции заслуженно известного Гарольда Пьюзи. Он умел, несмотря на свою сухую и нудную манеру изложения, увидеть за иссохшими костями животное из плоти и крови, жившее в давно ушедшую эпоху[81]. Собственное (хотя и происходящее от самого Ромера) объяснение причин, побудивших лопастеперых заполучить легкие и ноги, чтобы выйти на сушу, Пьюзи крепко вложил в мою студенческую голову. Я до сих пор вижу в нем смысл, несмотря на то, что в наши времена оно несколько менее в моде, чем во времена Ромера. И Ромер, и Пьюзи представляли себе ежегодные засухи, во время которых озера, пруды и речушки пересыхали, а в следующий паводок заполнялись снова. В этих условиях рыбы – водные обитатели – могли, однако, получать существенное преимущество от приобретенной способности жить на суше в короткие промежутки времени, когда им требовалось переползти из пересыхающего озера или пруда в более глубокий водоем, в котором можно дожить до следующего влажного сезона. Получается, что наши далекие предки не столько выходили на сушу, сколько пользовались ею как понтоном, по которому можно вернуться в воду. Многие современные нам животные поступают так же.
К сожалению, в предисловии к изложению своей теории Ромер показал, что девон был засушливым периодом. Поэтому, когда позднее выяснилось, что это не совсем так, появились сомнения и в справедливости самой теории Ромера. По‑моему, Ромеру просто следовало отказаться от этого введения, в любом случае претенциозного. В «Рассказе прародителя» я упоминал, что теория Ромера еще жизнеспособна, даже если климат в девоне был куда менее засушлив, чем предполагалось.
Как бы то ни было, вернемся к ископаемым. В позднем девоне – периоде, предшествовавшем каменноугольному – удалось обнаружить жалкие крохи. Однако эти крохи представляют собой волнующие следы «недостающих звеньев»: животных, которые продвинулись по пути от обычных в девонских морях лопастеперых рыб к амфибиям, заполнившим болота каменноугольного периода. В 1881 году в коллекции останков из Канады был обнаружен первый представитель этих животных, близких к лопастеперым – эустеноптерон. По‑видимому, это была рыба, охотившаяся у поверхности воды и, вопреки ранним предположениям, никогда не выходившая на сушу. Тем не менее она обладала анатомическими особенностями (кости черепа, зубы и, главное, плавники), характерными для амфибий, живших на 50 миллионов лет позднее. Вероятнее всего, эустеноптерон при помощи плавников плавал, а не ходил, однако кости плавников расположены характерным для тетрапод, то есть четвероногих (собирательное название наземных позвоночных), образом. В передней конечности единственная плечевая кость соединялась с двумя костями предплечья – лучевой и локтевой, а они, в свою очередь, присоединялись к нескольким небольшим костям, которые мы, настоящие тетраподы, называли бы пястью, запястьем и пальцами. Задние конечности были устроены аналогичным образом.
Затем, уже со стороны амфибий, где‑то на границе девона и карбона, то есть на 20 миллионов лет позднее, появляется ихтиостега (найдена в 1932 году в Гренландии). Кстати, пусть мысли о холоде и льдах вас не смущают. Во времена ихтиостеги Гренландия располагалась на экваторе. Первая реконструкция этого ископаемого животного была выполнена шведским палеонтологом Эриком Ярвиком в 1955 году. Он изображал ихтиостегу существенно ближе к сухопутным, чем считают современные эксперты. Согласно современной реконструкции Пера Альберга из Университета Уппсалы (родного университета Ярвика), ихтиостега большую часть времени проводила в воде, но совершала, вероятно, и недолгие вылазки на сушу. Тем не менее она куда сильнее напоминала саламандру, чем рыбу, и голова у нее была плоской, как у амфибий. В отличие от современных тетрапод, имеющих по пять пальцев на передних и задних конечностях (по крайней мере на эмбриональной стадии; во взрослом состоянии они могут часть пальцев утрачивать), у ихтиостеги было по семь пальцев. Похоже, что ранние тетраподы обладали большим простором для экспериментов в количестве фаланг и пальцев, чем мы теперь. По‑видимому, в какой‑то момент эмбриологические процессы зафиксировались на пяти пальцах, и был сделан шаг, после которого нам оказалось трудно повернуть вспять. Трудно, но не очень: встречаются же шестипалые кошки, а у людей бывает по шесть пальцев. Эти дополнительные пальцы возникают из‑за дупликации.
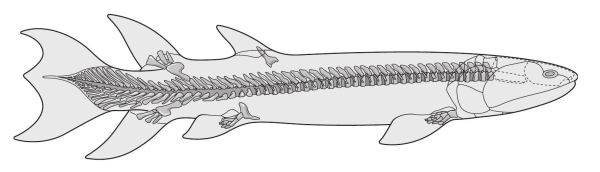
Эустеноптерон
Еще одно прекрасное ископаемое из тропической Гренландии, датируемое границей девона и карбона, было названо акантостегой. Как и ихтиостега, это животное имело плоский череп и конечности, характерные для тетрапод. Однако оно еще сильнее отличалось от того, что мы считаем стандартом: у акантостеги было восемь пальцев. Наибольший вклад в исследование акантостеги внесли Дженни Клак и Майкл Коутс из Кембриджского университета. Они считают, что акантостега, как и ихтиостега, была водоплавающим животным, но имела легкие. Устройство конечностей этого животного свидетельствует о том, что в случае необходимости оно могло перемещаться по суше практически так же хорошо, как и в воде. Выглядело же оно как гигантская саламандра. Возвращаясь к «рыбьей» стороне провала Ромера: пандерихтис, также живший в позднем девоне, был чуть более близок к амфибиям, чем эустеноптерон. Но если бы вы увидели его живьем, то назвали бы рыбой, а не саламандрой.
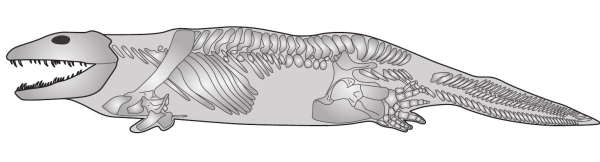
Ихтиостега
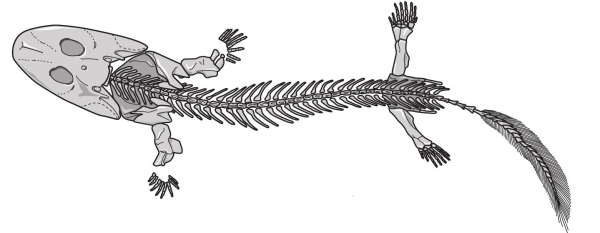
Акантостега
Таким образом, остается промежуток между амфибиеподобной рыбой пандерихтисом и рыбоподобной амфибией акантостегой. А где же звено между ними? Его поисками занималась группа из Университета Пенсильвании, в том числе Нил Шубин и Эдвард Дэшлер. Основываясь на результатах той экспедиции, Нил Шубин сделал серию блестящих замечаний об эволюции человека в своей книге «Внутренняя рыба». После долгого выбора места для поисков они отправились в горный район Канадской Арктики, содержавший породы позднего девона. Отправились – и нашли настоящее зоологическое сокровище. Тиктаалик! Это имя никогда не забудется[82]. По‑латыни – Tiktaalik, а на инуитском языке оно означает «крупная пресноводная рыба». В связи же с видовым названием тиктаалика (roseae) я могу рассказать поучительную историю о самом себе. Услышав это название и увидев фотографии (цветная вклейка 10), я представил себе классические девонские отложения, знаменитый «древний красный песчаник» графства Девон и города Петры (тот самый «город цвета красных роз», который «наполовину младше земных времен»[83]).

Пандерихтис
Как же я ошибся! На фотографиях розовый отчаянно преувеличен, а имя было выбрано в честь мецената, финансировавшего экспедицию в Арктику. Вскоре после этого открытия я обедал с доктором Дэшлером, и мне была оказана честь осмотреть находку. Признаюсь, мое зоологическое «я» (или моя «внутренняя рыба»?) были потрясены до потери дара речи. Я представил, что смотрю в лицо своего прямого предка. Как бы фантастично ни звучали мои слова, но эти не‑совсем‑розовые‑остатки были тем самым мертвым предком, который был всего наполовину младше земного времени.
Если бы вам довелось столкнуться с живым тиктааликом нос к носу, вы скорее всего сбежали бы: до того он напоминал крокодила. Голова крокодила на теле саламандры с рыбьим хвостом. У тиктаалика, в отличие от рыб, была шея, и он мог поворачивать голову. Со всех точек зрения тиктаалик представляет собой совершенное «недостающее звено» – совершенное потому, что он практически ровно наполовину рыба, наполовину – амфибия, а также потому, что тиктаалик теперь с нами. У нас есть его ископаемые остатки. Их можно увидеть, потрогать, даже попытаться почувствовать их древность – безрезультатно.
…и обратно
Выход из моря на сушу запустил цепочку преобразований в каждой сфере, от дыхания до размножения. Это был великий поход через биологическое пространство. Тем не менее впоследствии некоторые вполне успешные наземные животные отказывались от завоеванной земли и с необъяснимым упорством стремились обратно в воду. Тюлени и морские львы прошли эту дорогу наполовину. Они показывают нам, как могли выглядеть промежуточные звенья на пути к китам и дюгоням. Киты (включая мелких китов, которых мы называем дельфинами) и дюгони со своими близкими родственниками ламантинами совершенно оставили сушу и возвратились к жизни в море, как их далекие предки. Они вообще не выходят на берег, даже для размножения. Однако им так и не удалось выработать ничего похожего на жабры, утраченные предками, и поэтому они продолжают дышать атмосферным воздухом. К другим животным, возвращающимся в воду хотя бы на некоторое время, относятся: прудовики Lymnaeidae, водяные пауки Cybaeidae, жуки‑плавунцы Dytiscidae, крокодилы, каланы Lutra, морские змеи Hydrophiinae, куторы Neomys, галапагосские нелетающие бакланы Phalacrocorax harrisi, галапагосские морские игуаны Amblyrhynchus cristatus, водяные опоссумы Chironectes minimus, представители южноамериканских сумчатых, утконосы Ornithorhynchus anatinus, пингвины и водные черепахи.
Долгое время эволюция китов была для нас тайной за семью печатями, но в последние годы мы существенно приблизились к ее разгадке. Результаты, полученные молекулярной генетикой (см. главу 10), показывают, что наиболее близкими родичами китов из ныне живущих животных являются бегемоты, затем свиньи, а после – жвачные. Удивительно, что в результате генетического анализа гиппопотамы оказались в эволюционном отношении ближе к китам, чем к парнокопытным – свиньям и жвачным, которые внешне похожи на них гораздо сильнее. (Вот и еще один пример того, что внешнее сходство организмов далеко не всегда указывает на их родство. Мы уже отметили это несоответствие, когда говорили о латимериях, для которых люди являются более близкой родней, чем другие рыбы. Эта аномалия была порождена выходом на сушу наших далеких предков – выходом, который ускорил их эволюцию и в результате которого двоякодышащие и латимерии остались позади. Они больше похожи на других рыб, чем на нас, просто потому, что не покинули воду.) Бегемоты, по крайней мере отчасти, остались на суше и потому во многом похожи на своих далеких сухопутных родичей – жвачных, в то время как киты, их более близкие родичи, окончательно ушли в воду и изменились так сильно, что их родство с гиппопотамами укрылось ото всех ученых, кроме молекулярных генетиков. Как вышедшие на сушу далекие рыбьи предки будто бы отправились в космос или взлетели в небо на воздушном шаре, так и предки китов избавились от пут гравитации и свободно выплыли в пространство, освоившись в воде.
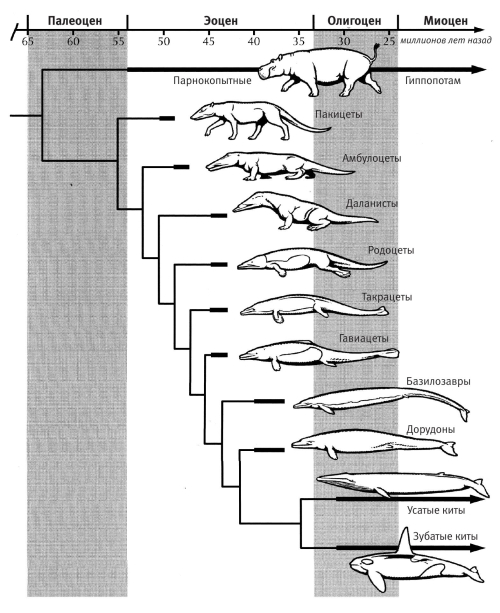
Эволюционный ряд, соединяющий наземных животных с китами и включающий многочисленные переходные формы, недавно обнаруженные в эоценовых отложениях Африки и Пакистана. Рисунок К. Буэля
В то же время палеонтологическая летопись китов, до сих пор практически полностью состоявшая из белых пятен, была в последние годы восстановлена – в основном благодаря находкам на территории современного Пакистана. История ископаемых китов настолько подробно описана в недавно вышедших книгах (в том числе в «Эволюция: о чем рассказывают ископаемые остатки» Дональда Протеро и «Почему эволюция истинна» Джерри Койна), что я решил подробно ее не пересказывать и ограничился диаграммой из книги Протеро.
Обратите внимание на осторожность, с которой нарисована схема. Любого автора подмывает (авторы старых книг часто уступали этому соблазну) изобразить последовательность ископаемых животных, соединенных стрелками, от древних к молодым. Однако никто не может утверждать наверняка, что Ambulocetus произошел от Pakicetus, а Basilosaurus – от Rodhocetus. Вместо этого осторожно предполагается, что киты произошли от животного, родственного Ambulocetus, похожего на него, возможно – именно от него. Изображенные на схеме ископаемые животные представляют собой разные стадии эволюции китов. Постепенная утрата задних конечностей, превращение передних конечностей в ласты, уплощение хвоста и его превращение в хвостовой плавник – вот некоторые изменения, поэтапный ход которых можно наблюдать в этой элегантной эволюционной последовательности.

Современный дюгонь (вверху) и древний дюгонь Pezosiren
Другая группа полностью водных млекопитающих – сирены (к ним относятся дюгони и ламантины) – имеет куда более неясную палеонтологическую летопись, однако недавно было найдено прекрасное «недостающее звено»: Pezosiren, «шагающий ламантин» с Ямайки[84]. Его ископаемые остатки имеют примерно такой же возраст, как и остатки Ambulocetus – жившего в эоцене «шагающего кита». В целом Pezosiren очень похож на современного ламантина или дюгоня. Отличаются они тем, что у Pezosiren были две пары ног, пригодных для ходьбы, а у его потомков вместо передних ног – плавники, а задних нет вовсе. На рисунке изображены два скелета: современного дюгоня и Pezosiren.
Существует множество свидетельств (включая самые важные – молекулярно‑генетические) того, что сирены – родственники слонов, точно так же как киты – родственники гиппопотамов. Тем не менее Pezosiren, вероятно, вел образ жизни, сходный с современными гиппопотамами – проводил большую часть времени в воде, пользуясь ногами и для ходьбы, и для плавания. Строение его черепа, однако, не оставляет сомнений в том, что перед нами представитель сирен. Неизвестно, был ли Pezosiren прямым предком современных дюгоней и ламантинов, но он, бесспорно, хорошо подходит на эту роль.
Когда книга готовилась к публикации, журнал «Нэйчер» напечатал статью об ископаемом животном[85], обнаруженном в канадской Арктике и закрывающем лакуну в эволюционной линии современных ластоногих (тюленей, морских львов и моржей). Найденный скелет (сохранилось около 65 % костей) был назван Puijila darwini. Он относится к раннемиоценовой эпохе (примерно двадцать миллионов лет назад). Это не очень давно: карта мира тогда была почти такой же, как сейчас. Следовательно, найденный ископаемый тюлень (или морской лев – их эволюционные пути тогда еще не разошлись) был арктическим животным, обитателем холодных вод. На основании имеющихся данных мы можем утверждать, что древний тюлень жил и ловил рыбу в пресных водоемах, подобно выдрам (за исключением калифорнийской морской выдры или калана – жителя океанического побережья). Среди современных тюленей большинство обитает в море, только байкальская нерпа населяет пресные воды. У Puijila еще не было ласт, но уже были лапы с перепонками. Вероятно, по земле он бегал как собака (не похоже на современных ластоногих), но проводил большую часть времени в воде, где плавал – также по‑собачьи, не прибегая ни к одному из двух способов плавания, впоследствии принятых тюленями и морскими львами. Puijila аккуратно закрывает пробел между сухопутными и водоплавающими существами в родословной ластоногих. Вот и еще одно пополнение в длинном списке звеньев, которые больше не являются недостающими.
Перейдем к другой группе животных, вернувшихся с суши обратно в воду. Пример поразителен еще и тем, что некоторые из них впоследствии вернулись на сушу еще раз! В отличие от китов и дюгоней, морские черепахи не полностью ушли в воду – они выходят на берег, чтобы отложить яйца. Как и все позвоночные, вернувшиеся в воду, они не смогли отказаться от дыхания атмосферным воздухом, но некоторые из них продвинулись в этом направлении несколько дальше китов. Эти водные черепахи, обладая двумя камерами на заднем конце тела, насыщенными кровеносными сосудами, способны извлекать кислород из воды. Одна из австралийских речных черепах получает большую часть кислорода для дыхания буквально, как без стеснения говорят австралийцы, через задницу.
Перед тем как приступить к обсуждению интересных эволюционных проблем, нам придется остановиться на занудных терминологических подробностях[86]. Связаны они с языком. Не случайно Бернард Шоу сказал: «Англия и Америка – это две страны, разделенные общим языком». Эта фраза с полным основанием относится к наименованиям черепах. В британском английском черепахи (turtles) – это те, кто живет в море. Для наземных черепах имеется особое слово (tortoises), свое наименование используется и для пресноводных черепах (terrapins). В американском английском все эти животные называются одним словом – черепаха. Поэтому для английского зоолога словосочетание «наземная черепаха» звучит дико, а для американского (и русского) – нормально. Иногда в американской литературе слово tortoises используется для обозначения отдельной группы черепах Testudinidae, а это современные наземные черепахи. А в британском английском этим словом называются все наземные черепахи, будь они представителями Testudinidae или другого семейства (мы вскоре увидим, что есть вымершие наземные черепахи, не принадлежащие к Testudinidae). Вот такая путаница. Но зоологи, на каком бы языке они ни говорили, называют обобщенно всех этих животных «черепаховые» или хелониевые (калька с латинского названия). Далее в тексте черепахи и хелониевые будут считаться синонимами.
|
|
|






