

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Топ:
Выпускная квалификационная работа: Основная часть ВКР, как правило, состоит из двух-трех глав, каждая из которых, в свою очередь...
Оценка эффективности инструментов коммуникационной политики: Внешние коммуникации - обмен информацией между организацией и её внешней средой...
Характеристика АТП и сварочно-жестяницкого участка: Транспорт в настоящее время является одной из важнейших отраслей народного...
Интересное:
Как мы говорим и как мы слушаем: общение можно сравнить с огромным зонтиком, под которым скрыто все...
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Берегоукрепление оползневых склонов: На прибрежных склонах основной причиной развития оползневых процессов является подмыв водами рек естественных склонов...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Частота смены поколений у ящериц P. sicula не превышает двух лет, поэтому эволюционные изменения, которые мы наблюдаем на острове Мрчара, произошли всего за восемнадцать‑девятнадцать поколений. Теперь задумайтесь: что если на протяжении трех‑четырех десятилетий следить за эволюцией бактерий? Бактерий, поколения которых сменяются не за годы, а в течение нескольких часов, даже минут? Бактерии преподносят эволюционисту и другой бесценный подарок: в некоторых случаях их можно на неограниченное время заморозить, а после оживить, и они продолжат размножаться как ни в чем не бывало. Таким образом, экспериментаторы могут составлять собственную «палеонтологическую летопись», делать мгновенный снимок состояния эволюционного процесса в любой его точке. Представьте: если бы мы могли вернуть к жизни «Люси», человеческого предка, чьи окаменелые остатки были найдены Дональдом Йохансоном, из глубокой заморозки и предоставить ей как представительнице своего вида возможность размножаться и эволюционировать! Именно это и сумел проделать с бактерией Escherichia coli Ричард Ленски и его коллеги из Мичиганского университета[60]. В наши дни многие исследования по плечу только большому коллективу ученых. Когда я буду рассказывать, что «Ленски сделал то‑то», меня следует понимать так: «Ленски со своими сотрудниками и студентами». Мы увидим, почему эксперименты Ленски так нервируют креационистов.
Эта работа – блистательная демонстрация эволюции.
Кишечная палочка E. coli – очень распространенный микроорганизм. Чрезвычайно распространенный. В любой момент по всему миру их примерно сто миллиардов миллиардов, причем около миллиарда из них находятся сейчас в вашем толстом кишечнике. Большинство кишечных палочек безвредно или даже приносит пользу, но иногда возникают и вредоносные штаммы. Столь частые эволюционные изменения не удивительны, если учесть огромную численность бактерий (хотя мутации происходят довольно редко). Предположим, что вероятность мутации любого гена в ходе единичного акта размножения составляет один на миллиард. Но бактерий настолько много, что любой ген наверняка мутирует ежедневно где‑нибудь в мире. Как говорит сам Ленски, «возможностей для эволюции – масса». Ричард Ленски и его коллеги воспользовались этими возможностями. Для понимания доказательной силы их экспериментов важны детали, поэтому я постараюсь доходчиво, но подробно изложить их.
|
|
Бактерии E. coli размножаются делением, то есть бесполым путем, и благодаря этому можно быстро и легко создать огромную популяцию генетически идентичных организмов. В 1988 году Ленски взял бактерии из одной такой популяции и поместил их в двенадцать идентичных колб, содержащих одну и ту же среду, основным питательным компонентом которой являлась глюкоза. Колбы с исходными популяциями бактерий помещались в шейкерный инкубатор, в котором подопытным было тепло и хорошо, и взбалтывались, чтобы бактерии были равномерно распределены в питательной среде. Первым бактериям из этих двенадцати колб было суждено основать двенадцать независимо эволюционирующих линий, которым предстояло существовать отдельно друг от друга свыше двадцати лет. Почти как двенадцать колен Израилевых, разве что у тех не было запрета смешиваться друг с другом.
Эти двенадцать «колен» E. coli не жили все время в одной и той же колбе: каждой линии ежедневно доставалась новая (вообразите двенадцать рядов колб, семь тысяч в каждом). Каждый день небольшим количеством жидкости из предыдущей колбы – ровно сотая часть содержимого – заражалась новая, богатой глюкозой среда. Численность популяции бактерий в новой колбе начинала стремительно расти, однако через сутки темп роста всегда выходил на плато, потому что запас пищи иссякал. Другими словами, сначала численность популяции в колбе резко возрастала, затем достигала плато, и в этот момент из популяции отбиралась новая проба, которую пересаживали в новую колбу, и цикл начинался заново. Таким образом, бактерии проходили через каждодневные циклы изобилия, сменяющегося голодом, от которого спасалась счастливая сотая часть, которую, как в стеклянном Ноевом ковчеге, переносили в новый, пусть временный, глюкозный рай. Замечательные, чудесные условия для эволюции. Немаловажно, что эксперимент шел параллельно на двенадцати линиях.
|
|
Ленски и его команда продолжали повторять эти операции ежедневно на протяжении двадцати с лишним лет. С начала эксперимента сменилось примерно 7000 «поколений колб» и около 45000 поколений бактерий (в день сменяется шесть‑семь поколений). Если бы мы проследили столько же поколений назад в истории человека, то оказались бы примерно за миллион лет до сегодняшнего дня, во времена Homo erectus – что, впрочем, по эволюционным меркам не так уж много. Задумайтесь на секунду – если уж за время, эквивалентное миллиону лет по меркам млекопитающих, Ленски наблюдает эволюционные изменения, сколько изменений должно было произойти за сто миллионов лет реальной эволюции млекопитающих? А ведь сто миллионов лет – это не так уж много с точки зрения геологии.
Помимо основного эволюционного эксперимента, группа Ленски использовала культуру кишечной палочки для нескольких побочных опытов, в одном из которых, например, через две тысячи поколений бактерий глюкозу заменили другим сахаром – мальтозой (мы остановимся на основной работе, в которой использовалась глюкозная среда). Через равные интервалы времени ученые отбирали пробу бактерий каждого из «колен», чтобы понять, как идет эволюционный процесс. Кроме того, они замораживали взятые образцы, чтобы получить своеобразную палеонтологическую летопись бактериальной эволюции.
Трудно переоценить замысел этих экспериментов. Приведу замечательный пример планирования. Помните, я говорил, что бактерии во всех двенадцати колбах происходили из одного клона и были, таким образом, вначале генетически идентичными? На самом деле это не совсем так. Незадолго до начала этой работы в лаборатории Ленски изучал ген ara, встречающийся в двух вариантах: Ara + и Ara‑. Вы не сможете различить их, пока не посеете культуру бактерий в чашку с агаром, содержащим питательную среду, сахар арабинозу и краску тетразоль. Посев – распространенный в микробиологии прием, при котором на дно чашки, покрытое тонким слоем застывшего агарового геля, наносится небольшое количество жидкости, содержащей бактерии, затем чашка инкубируется. Колонии бактерий вырастают из упавших на агар капель в виде концентрических колец, питаясь находящимися в агаре питательными веществами. Если среда содержит арабинозу и краску‑индикатор, то различия между двумя генами становятся очевидными: будто кто‑то нагрел невидимые чернила. Колонии, содержащие Ara‑, становятся красными, Ara + – белыми. Ленски решил, что это различие удобно в качестве метки, и сделал шесть «колен» Ara +, шесть – Ara‑. Использовали их исследователи, например, для того, чтобы проверять собственные лабораторные методы. Производя ежедневный ритуал посева, они тщательно следили за тем, чтобы попеременно работать с линиями Ara + и Ara‑. Таким образом, если ученые допустили бы ошибку, то она проявилась бы на красно‑белом тесте. (Гениально, не правда ли? И скрупулезно. Хорошему ученому приходится сочетать эти качества.)
|
|
Но забудем пока о гене ara. Во всем остальном клоны‑основатели были идентичными. Никаких других различий между Ara + и Ara‑ обнаружить не удалось, так что ими действительно можно пользоваться просто как цветными маркерами, вроде колец на птичьих лапах.
Прекрасно. У нас есть двенадцать бактериальных «колен», мчащихся сквозь собственное «геологическое время», переживая циклы изобилия и голода. Останутся ли они такими же, как их предки? Или начнут изменяться? А если начнется эволюция, то будут ли все двенадцать «колен» изменяться одинаково? Как я уже говорил, питательная среда содержала глюкозу.
Не будучи единственным источником пищи, она, тем не менее, являлась ограничивающим рост колонии ресурсом. Причиной прекращения взрывного роста и выхода на плато день за днем было именно истощение запасов глюкозы. Другими словами, если бы экспериментаторы клали в колбы чуть больше глюкозы, то плато в конце каждого дня было бы выше. Если экспериментаторы решили бы добавить вторую дозу глюкозы после достижения плато, началась бы следующая стадия быстрого роста популяции – до нового плато.
|
|
В таких условиях, согласно дарвиновской теории, следует ожидать, что если произойдет мутация, помогающая отдельной бактерии использовать глюкозу более эффективно, то естественный отбор ее поддержит и мутация распространится в данной колбе, потому что мутантные бактерии будут размножаться быстрее, чем не‑мутантные. Поэтому в следующую колбу бактерии‑мутанты попадут в повышенной пропорции, а затем, переходя из колбы в колбу, довольно быстро добьются монополии в своем «колене». Именно это и случилось во всех двенадцати «коленах» – со сменой поколений произошло полное замещение предковых популяций бактериями, способными лучше усваивать глюкозу. Но, что самое поразительное, они добились лучшего усвоения глюкозы различными путями. То есть разные линии бактерий выработали различные наборы мутаций.
Как ученым удалось это узнать? Сравнивая уровень «приспособленности» бактерий в каждой линии с замороженными «ископаемыми» – предковыми популяциями. Напомню, «ископаемые» в данном случае – это замороженные бактерии, которые после размораживания способны нормально жить и размножаться. Как же, спросите вы, Ленски и коллеги сравнивали приспособленность бактерий с их «ископаемыми» предками? Весьма изящным образом. Образец эволюционировавшей линии отбирался и помещался в новую колбу. Туда же помещалось равное количество размороженной культуры. Разумеется, контакт с бактериями основных двенадцати линий сразу же исключался. Используемые для этого побочного опыта образцы в эксперименте участия не принимали.
Итак, перед нами колба с двумя конкурирующими культурами, «современной» и «ископаемой», и мы желаем знать, какая из линий окажется успешнее и достигнет наибольшей численности. Помните про «цветное кодирование» при помощи гена ara? Что нам потребуется, чтобы сравнить конкурентоспособность «современных» бактерий, например, из линии № 5 с «ископаемым» предком? Допустим, линия № 5 имеет ген Ara +. Тогда возьмем вариант Ara‑ предковой популяции, разморозим и добавим в «соревновательную» колбу. Если же линия № 5 несла вариант Ara‑, то ее следовало бы смешивать с предковой популяцией Ara +. Из предыдущих работ группы Ленски было известно, что сами по себе гены Ara + и Ara‑ не оказывают действия на приспособленность бактериальной популяции. Поэтому исследователи могли использовать цветные маркеры, чтобы сравнить конкурентоспособность каждого эволюционирующего «колена» с «ископаемыми» предками. Для этого надо было высадить каплю смеси из «соревновательной» колбы в чашку Петри и сосчитать, сколько колоний будут белыми, а сколько – красными.
|
|

Эксперимент Ленски. Размер бактериальной клетки в одной линии
Выше я упоминал, что приспособленность бактериальных культур с проходящими поколениями повышалась по сравнению с их предками. Все двенадцать «колен» стали успешнее выживать в условиях дефицита глюкозы. Повышение приспособленности могло идти несколькими путями. Скорость размножения бактерий всех «колен» росла от колбы к колбе, и средний размер бактериальной клетки увеличивался (см. иллюстрацию). Точками отмечены реальные данные, кривая построена при помощи численной аппроксимации. Наилучшим приближением к наблюдаемому набору данных является гипербола. Всегда есть шанс, что математическая функция порядка более высокого, чем гипербола, обеспечит лучшее приближение к набору наблюдаемых данных, но и гипербола достаточно хороша. Биологи часто прибегают к статистическому анализу данных и вписыванию приближенной кривой в наблюдаемые данные, но, в отличие от физиков, мало когда сталкиваются с настолько точным совпадением. Обычно наши данные куда запутаннее. Биолог может получить гладкую кривую только при наличии огромного количества тщательно собранных данных. Опыт Ленски прошел как по учебнику.
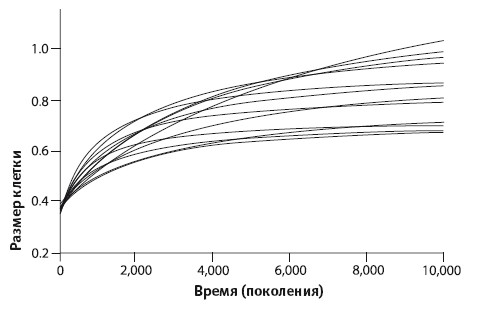
Эксперимент Ленски. Размер бактериальной клетки в двенадцати линиях
Как видите, основной прирост среднего размера бактериальной клетки произошел в течение первых двух тысяч поколений. Если все двенадцать «колен» увеличили в ходе эволюции средний размер бактериальной клетки, происходило ли это одним и тем же способом? Ответ – нет, и это второй интересный результат данной работы. График относится только к одной из двенадцати линий. Теперь посмотрим на соответствующие ему гиперболы для остальных «колен»: как велик разброс! Да, все они рано или поздно выходят на плато, но самое высокое плато почти вдвое выше самого низкого. Формой они также различаются: кривая, достигающая наибольшего значения после 10000 поколений, сначала растет медленнее многих других и «перехватывает инициативу» только после 7000 поколений. Не путайте эти плато с ежедневными плато численности популяции в каждой колбе: сейчас мы рассматриваем кривые в эволюционном масштабе времени, измеряемом поколениями колб, а не в индивидуальном масштабе, измеряемом часами существования в одной колбе.
Итак, увеличение размеров клетки, похоже, помогает бактериям выжить в условиях постоянного чередования изобилия и голода. Я не буду рассуждать о причинах этого (их может быть много), однако этим путем пошли все двенадцать линий. Правда, существует множество способов стать больше, что предполагает разные наборы мутаций, и, похоже, разные линии выработали в процессе эволюции разные подходы к увеличению размеров клетки. Это очень интересно. Еще интереснее то, что в некоторых случаях способ увеличения размера клетки совпал у двух линий. Ленски и другая группа его сотрудников исследовали это явление, взяв две клеточные линии Ara +1 и Ara‑ 1, которые в течение жизни 20000 поколений шли по одной и той же эволюционной траектории, и проанализировали их ДНК. Результат оказался ошеломляющим: в клетках этих двух линий изменилась активность (уровень экспрессии) 59 генов, и в обеих линиях уровень экспрессии всех 59 генов изменился в одном и том же направлении. Без помощи естественного отбора вероятность такого параллельного, независимого изменения активности 59 генов в двух линиях была бы предельно низка. Именно о таких вещах креационисты любят говорить, что те попросту не могут произойти, поскольку вероятность совпадения ничтожна. Но в действительности это произошло! Объяснение простое: никакой случайности здесь нет. Естественный отбор постепенно отдавал предпочтение одним и тем же полезным изменениям в обеих линиях.
Плавность кривой возрастания размера клетки с ходом поколений наводит на мысль, что улучшения в клетке происходят постепенно. Но не слишком ли она плавная? Не следует ли ожидать ступеней, временных остановок на пути эволюции, пока популяция ждет следующей увеличивающей приспособленность мутации? Не обязательно. Это зависит от множества факторов, в том числе от количества вовлеченных мутаций, силы влияния определенной мутации на размер клетки, негенетических факторов, влияющих на ее размер, частоты отбора проб. Но если мы посмотрим на график роста приспособленности (а не увеличения размера клетки), то увидим несколько иную картину, которую в принципе можно интерпретировать как последовательность ступеней (см. иллюстрацию ниже). Выше я упомянул, что всегда есть вероятность существования функции, более точно аппроксимирующей набор данных, чем та, которую мы выбрали. Математики называют любую такую функцию «моделью». Наши данные можно соотнести с гиперболической моделью, однако ступенчатая модель аппроксимирует их еще точнее.
Соответствие, однако, не столь близкое, как в случае с графиком роста клеток. Ни в одном из случаев нельзя доказать, что данные полностью соответствуют избранной модели, поскольку это невозможно. Однако данные по крайней мере не противоречат предположению, что мы наблюдаем эволюционные изменения, вызванные постепенным накоплением мутаций[61].
То, что мы увидели – прекрасный пример эволюции в действии. Эволюции, происходящей прямо перед нашими глазами, эволюции тщательно документированной, описанной сравнением двенадцати независимых линий между собой и с «живыми ископаемыми» – предками, вернувшимися, словно из глубин тысячелетий, из морозильника.
Теперь мы готовы к тому, чтобы познакомиться с еще более интригующими подробностями. До сих пор мы говорили, что все двенадцать «колен» развивались примерно в одном и том же направлении, постепенно приспосабливаясь к условиям роста, и различались только одним: кто‑то эволюционировал быстрее, кто‑то медленнее. Однако долгий эксперимент породил одно очень важное исключение. Вскоре после поколения № 33000 произошло нечто удивительное: одна из двенадцати линий, Ara ‑3, внезапно словно взбесилась. Взгляните на рисунок: на оси ординат отложена оптическая плотность, или «мутность» раствора, отражающая численность бактерий в колбе. Жидкость мутнеет просто потому, что в ней много бактерий; прозрачность жидкости может быть точно измерена, и результат мы можем использовать как показатель плотности популяции.
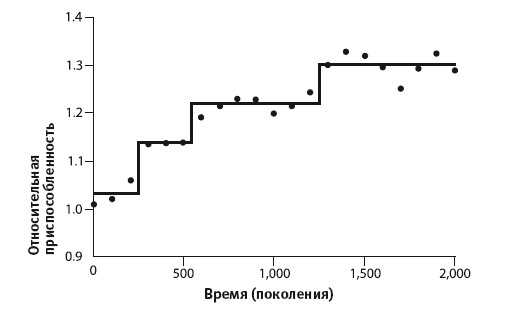
Эксперимент Ленски. Рост приспособленности
Если вы взглянете на рисунок, то заметите, что до поколения № 33000 показатель численности бактерий линии Ara ‑3 колебался в районе 0,04, мало отличаясь от остальных линий. Затем, сразу после поколения № 33100, плотность популяции Ara ‑3 начала чрезвычайно быстро, почти вертикально, расти. Среднее значение выросло почти до 0,25, то есть в шесть раз (!). Для этой популяции наступил «золотой век». Спустя всего несколько дней типичная высота плато, на которой стабилизировалась численность бактерий в популяции, выросла в шесть раз по сравнению с недавно еще таким постоянным значением, которое демонстрировали остальные линии. Возросшая высота плато впоследствии достигалась всеми поколениями линии Ara ‑3, и никогда – бактериями других линий. Все выглядело так, как будто линии Ara ‑3 досталась большая, чем другим, порция глюкозы. Однако никто этим бактериям лишней глюкозы не давал: во все колбы помещалось одинаковое, тщательно отмеренное ее количество.
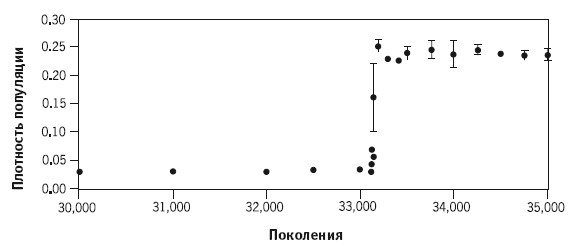
Эксперимент Ленски. Плотность популяции
Что же случилось с линией Ara ‑3? Ленски и двое его коллег потратили некоторое время на выяснение этого, и их усилия окупились сторицей. Произошло нечто замечательное.
Помните, я отметил, что ресурсом, ограничивающим рост популяции, является глюкоза, и линия, которая нашла бы более эффективный способ ее использования, получила бы эволюционное преимущество? Именно это и происходило во всех двенадцати «коленах». Но я говорил также о том, что глюкоза была не единственным питательным веществом в культуральной среде. Кроме глюкозы там был цитрат (лимонная кислота). Цитрата в среде было много, однако бактерия E. coli не способна его усваивать – по крайней мере, не в среде, в которой присутствует растворенный кислород (как это было в колбах Ленски). Но стоило бы хоть одной бактерии научиться усваивать цитрат, перед ней открылось бы настоящее Эльдорадо. Именно это произошло с линией Ara ‑3. Эта линия, и только она, внезапно обрела способность усваивать не только глюкозу, но и цитрат. Таким образом, количество пищи в колбе резко возросло, и, соответственно, резко вырос уровень плато, на котором стабилизируется популяция в конце каждого суточного цикла.
Выяснив, что случилось с загадочной Ara ‑3, Ленски и его коллеги продолжили работу, задавшись следующим важным вопросом. Была ли эта неожиданно появившаяся способность вызвана всего одной редкой мутацией (настолько редкой, что из двенадцати линий повезло только одной)? Была ли она, в сущности, такой же мутационной ступенью, как и другие мутации, обеспечившие ступенчатый рост приспособленности на графике выше? Ленски счел это маловероятным. Он предложил следующее объяснение. Зная средний темп мутаций в каждом гене бактерии, он вычислил, что время жизни 30000 поколений – это срок, достаточный для того, чтобы каждый ген мутировал хотя бы раз в каждой из двенадцати клеточных линий. Следовательно, маловероятно, что Ara ‑3 выделилась из общей массы благодаря редкости мутации. Будь это единичная мутация, ее бы открыли и в других линиях.
Но есть еще одна возможность, по крайней мере, теоретическая. Что если для того, чтобы питаться цитратом, бактериям необходима не одна мутация, а две, три, четыре? Мы не говорим сейчас о простом случае, когда эффекты двух мутаций суммируются. Если бы это было так, бактерии могли бы приобрести эти мутации порознь, в любом порядке. Любая из этих мутаций отдельно приближала бы нас на шажок к цели, предоставляла бы клетке способность частично усваивать цитрат, пусть не так эффективно, как две мутации вместе. Мутации такого типа мы уже обсуждали, когда рассматривали механизм увеличения размера клетки. Однако в данном случае приобретение способности питаться цитратом не было бы такой редкостью, и линия Ara ‑3 не оказалась бы уникальной. Редкость цитратного метаболизма обязывает нас искать что‑то более похожее на столь любимую креационистами «несократимую сложность». Может существовать биохимический путь, состоящий из двух реакций, причем в ходе первой реакции образуется необходимый исходный материал для второй, а для получения конечного продукта необходимы обе реакции. Для создания такого механизма потребовались бы две мутации (назовем их А и Б), катализирующие две реакции. При этом только две мутации вместе дают бактерии преимущество, а по отдельности они бесполезны и потому не могут быть поддержаны отбором. Такое двойное событие, без сомнения, будет достаточно редким для того, чтобы произойти только в одном из двенадцати случаев.
Однако пока это гипотеза. Была ли у группы Ленски возможность экспериментально выяснить, что произошло? Ученые могли бы сделать ряд очень важных шагов в этом направлении, используя размороженные «ископаемые». Повторим основные положения нашей гипотезы. В некоторый момент в геноме Ara ‑3 произошла мутация А. Она не имела заметного эффекта, поскольку мутация Б, необходимая для цепочки биохимических реакций, отсутствовала. Мутация Б может с одинаковой вероятностью возникнуть в любой линии. Более того, вполне возможно, что она там возникала! Но мутация Б бесполезна – она не дает абсолютно никакого положительного эффекта, если линия предварительно не обзавелась мутацией А. Так получилось, что подготовлена таким образом была только линия Ara ‑3.
Ленски мог бы даже сформулировать гипотезу в форме проверяемого предсказания – и, пожалуй, имеет смысл так ее и сформулировать, потому что это и есть самое настоящее предсказание, хоть оно и относится к прошлому. Вот как я бы сформулировал ее на месте Ленски:
Я разморожу «ископаемые» образцы линии Ara ‑3 разного, сознательно выбранного возраста. Затем я дам «воскрешенным» бактериям возможность эволюционировать в тех же условиях, что и в основном эксперименте, но в полной изоляции от других. Теперь – собственно предсказание. Способность усваивать цитрат приобретут только линии размороженных бактерий, которые были заморожены до определенного, критически важного поколения основного эксперимента. Так мы сможем установить, когда в линии появилась мутация А.
Вы, вероятно, обрадуетесь, когда узнаете, что именно к этому результату пришел студент Ленски Захария Блаунт. Он провел изматывающую серию экспериментов с сорока триллионами (40.000.000.000.000) клеток E. Coli из разных поколений и получил результат, полностью соответствующий такому предсказанию. Чудо произошло примерно в поколении № 20000. Замороженные бактерии из последующих поколений демонстрировали повышенную вероятность появления способности к усвоению цитрата, чего не показывали бактерии из более ранних поколений. В полном согласии с нашей гипотезой, клоны после поколения № 20000 были подготовлены к восприятию мутации Б. Все «воскрешенные» бактерии, относившиеся к поколениям после № 20000, имели одинаковую вероятность появления способности к усвоению цитрата (прежде эта вероятность не была повышена). До этого момента линия Ara ‑3 ничем не отличалась от прочих. Бактерии еще не имели мутации А. Однако после поколения № 20000 только линия Ara ‑3 могла воспользоваться позитивным эффектом мутации Б. В жизни ученого есть моменты, когда он испытывает истинное наслаждение от результата своего труда. Это, безусловно, был один из них.
Работы Ленски демонстрируют нам в масштабе микрокосма и условиях лаборатории многократно ускоренные процессы эволюции путем естественного отбора. Мы наблюдаем: случайную мутацию, за которой следует неслучайный отбор; независимые пути адаптации к одним и тем же внешним условиям у разных популяций; эволюционные изменения за счет добавления успешных мутаций к существующим; зависимость эффектов некоторых генов от присутствия других генов. И все это произошло за время, составляющее ничтожную долю срока, который обычно требуется для эволюционных изменений в природе.
У этой истории триумфа есть комическое продолжение. Креационисты ее ненавидят. Ведь здесь заметна не только эволюция в действии, не только попадание новой информации в геном безо всякого вмешательства разумного Творца (эту возможность им сказано было отрицать при любой возможности; я говорю «им было сказано», поскольку большинство креационистов плохо представляет себе, что такое информация), не только мощь естественного отбора, позволяющая ему составлять комбинации генов, которые, исходя из столь любимых креационистами вероятностных вычислений, статистически невозможны. Хуже всего для креационистов то, что эксперимент Ленски не оставляет камня на камне от постулата о «несократимой сложности». Так что нет ничего удивительного в том, что они озабочены исследованиями Ленски.
Эндрю Шлэфли, креационист и редактор «Консервапедии», печально известного аналога «Википедии», написал Ричарду Ленски и потребовал доступа к его исходным данным, высказав сомнение в их достоверности. В принципе, Ленски вовсе не был обязан отвечать, но тем не менее вежливо посоветовал Шлэфли прежде прочитать его статью, а уж затем критиковать работу. Кроме того, он отметил, что основные его данные – это замороженные бактериальные культуры, которые кто угодно может исследовать, чтобы проверить достоверность полученных результатов. Он с радостью готов предоставить эти культуры любому микробиологу, который будет в состоянии с ними работать – поскольку в неопытных руках они могут быть весьма опасными. Ленски подробно перечислил требования, предъявляемые к микробиологу. Любой читатель может представить себе, с каким удовольствием он это сделал, отлично зная, что Шлэфли – адвокат, а не ученый, и поэтому просто не поймет половину сказанного, не говоря уже о том, что не проведет необходимые исследования и статистический анализ результатов. Черту под историей подвел известный ученый и блогер Пи Зет Майерс[62], комментарий которого начинался словами «В который уже раз Ричард Ленски утер нос недотепам и дуракам из „Консервапедии “, и, боже мой, насколько же он круче».
Проведенные Ленски эксперименты, особенно гениальная методика заморозки «ископаемых образцов», показывают способность естественного отбора вызывать эволюционные изменения за промежутки времени, которые мы в состоянии оценить и понять, то есть сопоставимые со сроком нашей жизни. Однако бактерии представляют нам и другие впечатляющие примеры, пусть и не так детально изученные. Многие штаммы бактерий удивительно быстро выработали устойчивость к антибиотикам. Ведь первый антибиотик – пенициллин – был героически разработан Флори и Чейном во время Второй мировой войны, то есть не так давно. С тех пор регулярно появляются новые антибиотики, и бактерии выработали устойчивость почти к каждому из них. На сегодняшний день одним из самых зловещих примеров является MRSA (устойчивый к метициллину Staphylococcus aureus), сделавший многие больницы весьма опасными для посещения местами. Другой нешуточной угрозой является микроорганизм Clostridium difficile. И в этом случае мы наблюдаем получение штаммами, обладающими устойчивостью к антибиотикам, эволюционного преимущества в ходе естественного отбора, однако здесь добавляется еще один фактор.
При длительном лечении антибиотиками гибнут практически все микроорганизмы в кишечнике человека – как патогенные, так и полезные. Однако C. difficile, устойчивый к большинству антибиотиков, получает огромные преимущества на фоне отсутствия микроорганизмов, с которыми он в нормальных условиях конкурировал бы («враг моего врага – мой друг»).
Меня неприятно удивила брошюра в приемной моего врача, предупреждающая об опасности неоконченного курса приема антибиотиков. Такая опасность существует, и предупреждать о ней правильно, но меня озадачила названная в брошюре причина. Там объяснялось, что бактерии «умны» и способны «учиться» справляться с антибиотиками. Вероятно, авторам брошюры показалось, что объяснить природу устойчивости микроорганизмов к антибиотикам обычному человеку будет проще, если указание на естественный отбор заменить словами об обучении. Однако разговоры о том, что бактерии «умны» и способны к «обучению», сбивают пациентов с толку и, главное, не убеждают в необходимости продолжать прием антибиотиков так долго, как это предписано врачом. И глупцу понятно, что странно рассуждать об «уме» бактерий. Кроме того, если бактерии были бы умны, как преждевременное прекращение приема антибиотиков повлияло бы на их когнитивные способности? Зато стоит нам вспомнить о естественном отборе, и разговор немедленно обретает смысл.
Действие антибиотиков, как и любого яда, зависит от дозировки: большая доза убьет все бактерии, средняя – не все, небольшая – ни одной. Если среди бактерий есть генетическая изменчивость, например, если одни из них устойчивее к антибиотикам, чем другие, то промежуточная доза будет стимулировать естественный отбор по признаку устойчивости к антибиотикам. Когда врач велит вам продолжать прием лекарства, он делает это для того, чтобы повысить вероятность гибели всех бактерий, не оставляя в живых устойчивых или полуустойчивых мутантов. Будь мы ближе знакомы с теорией Дарвина, мы смогли бы раньше осознать опасность появления устойчивых к антибиотикам микроорганизмов. К сожалению, брошюры вроде той, которую я видел в приемной своего врача, ничуть не способствуют просвещению. Очень жаль, что упущена замечательная возможность рассказать людям о могуществе естественного отбора.
Гуппи
Мой коллега доктор Джон Эндлер, недавно переехавший из Северной Америки в Эксетер, рассказал мне замечательную, хоть и невеселую, историю. Эндлер, будучи в Америке, летел внутренним рейсом, и попутчик спросил его, чем он занимается. Эндлер ответил, что он – профессор биологии, изучает популяции диких гуппи Тринидада. Пассажир заинтересовался и принялся расспрашивать Эндлера. Восхищенный элегантностью теории, на которой, очевидно, основывались эксперименты, попутчик спросил Эндлера, что это за теория и кто ее автор. Тогда‑то Эндлер и обрушил на голову собеседнику взрывоопасную информацию: «Это дарвиновская теория эволюции путем естественного отбора». Вдруг поведение собеседника совершенно изменилось. Он побагровел, резко отвернулся и прекратил всякое общение, отказавшись от продолжения прекрасной беседы. Воистину прекрасной: Эндлер рассказал мне, что перед этим конфузом сосед «задал несколько отличных вопросов, указывающих на то, что он с удовольствием и пониманием участвовал в беседе. Поистине горько».
Эксперименты, о которых Эндлер рассказывал своему предвзятому попутчику[63], просты и элегантны и прекрасно иллюстрируют скорость, с которой может идти естественный отбор. Мне кажется, что работам Эндлера на страницах этой книги самое место, особенно учитывая, что он – автор труда «Естественный отбор в дикой природе» – лучшей на сегодняшний день книги с примерами подобных исследований и описанием методов.
Гуппи – широко известная пресноводная аквариумная рыба. Самцы гуппи (как и фазанов, с которыми мы встречались в главе 3) окрашены ярче самок. Аквариумисты поддержали усилия природы и вывели еще более ярких рыб. Эндлер изучал диких гуппи Poecilia reticulata, обитающих в горных реках Тринидада, Тобаго и Венесуэлы. Он заметил, что географически обособленные популяции заметно отличаются друг от друга: в некоторых самцы были окрашены во все цвета радуги и по яркости приближались к выращенным в аквариумах. Эндлер предположил, что предки этих самцов отбирались самками в зависимости от окраски, – то же самое происходило у фазанов. В других районах самцы гуппи, напротив, были окрашены гораздо скромнее, хотя все же ярче самок. Они, как и самки, обладали камуфляжной окраской, соответствующей каменистому дну ручьев, в которых они жили. Сравнивая численные показатели популяций из различных районов Венесуэлы и Тринидада, Эндлер показал, что в тех ручьях, где наблюдалась блеклая окраска самцов, гуппи активнее поедались хищниками. В ручьях же, где хищников меньше, самцы окрашены броско, число цветных пятен на их теле больше, пятна крупнее и ярче. При минимуме хищников самцы могли свободно развивать окраску, привлекающую самок, а там, где хищников больше, лучше поостеречься. Во всех популяциях присутствует эволюционное давление самок на самцов, заставляющее последних вырабатывать более привлекательную окраску, вне зависимости от наличия хищников и давления с их стороны. Эволюция, как всегда, находит компромисс между противоречивыми давлениями отбора. Эндлеру удалось не только проанализировать, как модель компромисса варьируется от популяции к популяции, но даже перейти к экспериментам.
Представьте себе, что вы хотите провести идеальный эксперимент и продемонстрировать эволюцию маскирующей окраски. Что вы стали бы делать? Камуфляж животных напоминает фон, на котором им приходится существовать. Можно ли провести эксперимент, в рамках которого животные будут активно приспосабливаться к среде, которую создадите для них вы? Или – еще лучше – создать два фона, по одному на каждую из двух изолированных популяций? В идеале следует провести нечто аналогичное эксперименту с двумя линиями кукурузы, описанному в главе 3. Но в нашем новом эксперименте отбор будет вести не человек, а хищники и самки гуппи. Единственное, чем будут различаться две экспериментальных популяции, – это фон, на котором они будут жить.
Возьмем какое‑нибудь животное с камуфляжной окраской – например, насекомое – и рассадим представителей одного вида по разным вольерам (прудам, клеткам и так далее) с разноцветным фоном. Например, можно сделать для одних вольеров зеленый фон (лес), для других – порыжелый (пустыня). Поместив экспериментальных животных в определенную среду, предоставим им жить и размножаться в течение жизни стольких поколений, сколько мы можем себе позволить, а затем вернемся и посмотрим, выработалось ли у них сходство с фоном, зеленым или красным соответственно. Естественно, ожидать такого результата мы можем, только если поместим в тот же вольер хищников. Значит, давайте запустим внутрь, скажем, хамелеона. Во все вольеры? Нет, это же эксперимент. Значит, нам нужен контроль – и мы поместим хищников в половину красных и половину зеленых вольеров. Цель нашего эксперимента – проверить предположение о том, что в вольерах с хищниками будет происходить эволюция в направлении камуфляжной окраски (зеленой ил<
|
|
|

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...

Архитектура электронного правительства: Единая архитектура – это методологический подход при создании системы управления государства, который строится...

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!