У круглоротых в спинном мозге намечается дифференциация мотонейронов на популяции, иннервирующие отдельные миомеры. Каждая такая популяция включает в себя группы первичных (крупных) и вторичных (мелких) мотонейронов, иннервирующих внутри отдельного миомера различные зоны (дорсальную или вентральную) и типы мышц. Первичные и вторичные мотонейроны образуют множественные контакты (от 3 до 8) на волокнах быстрых "белых" мышц - полинейронная иннервация. Мононейронная иннервация, осуществляемая только первичными мотонейронами, характерна для медленных "красных" мышц. Однако у круглоротых нет обособления отдельных популяций мотонейроров, иннервирующих разные типы мышц в пределах сегмента.
У хрящевых рыб дифференциация белого и серого вещества в спинном мозге нечеткая (рис. 20). В вентральных рогах имеются три группы клеток: дорсомедиальная, вентролатеральная и центральная. Для мотонейронов характерна большая вариабельность формы и размеров, а также тенденция к тесному расположению их тел и отростков. Особенность мотонейронов рыб состоит в том, что они увеличиваются в размерах в течение всей жизни организма (в отличие от нейронов других позвоночных).
У акул и скатов мотонейроны, иннервирующие красные и "белые мышцы миомеров, уже пространственно разграничены (рис. 20, в, г). Красные мышцы иннервируются мелкими мотонейронами, сосредоточенными в латеральной части мотонейронного пула, белые - крупными мотонейронами, занимающими медиальные районы. Более того, аксоны мотонейронов, иннервирующие
разные участки миомеров, имеют разный диаметр и проходят в разных участках вентральных корешков.
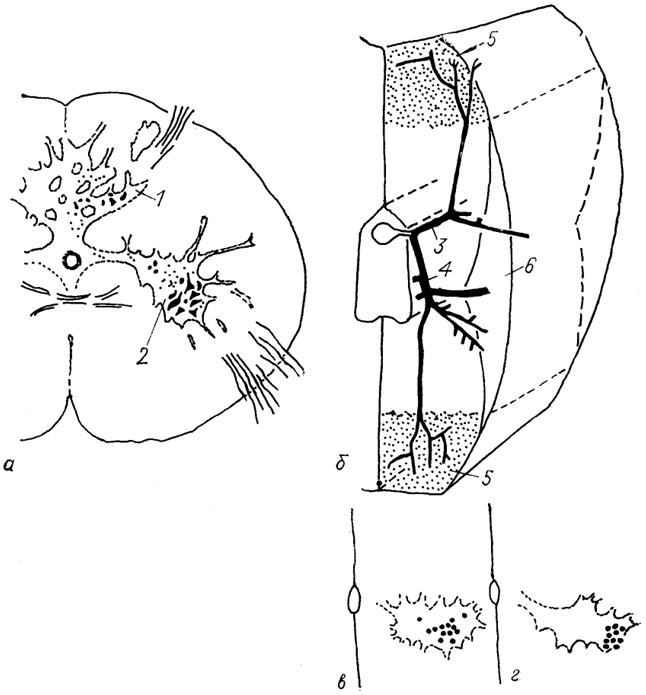
Рис. 20. Цитоархитектоника спинного мозга (а), ветвление вентрального корешка спинномозгового нерва (б) и локализация мотонейронов, иннервирующих "красные" (в) и "белые" (г) мышцы у акулы (Mos, Willamson, 1986).
1, 2 - задние (1) и передние (2) рога серого вещества; 3, 4 - дорсальная (3) и вентральная (4) ветви вентрального корешка спинномозгового нерва; 5, 6 - зона "белых" (5) и "красных" (6) мышц миотома.
Мотонейроны костистых рыб расположены в вентральных рогах серого вещества спинного мозга и представлены двумя популяциями. Первую популяцию составляют крупные первичные мотонейроны, локализованные исключительно в дорсальной части мотонейронной колонны. Их сильно развитые дендриты проникают в белое вещество и часто достигают противоположной стороны спинного мозга. Таких мотонейронов немного: например, у золотой рыбки на 1/2 сегмента приходится 8-12
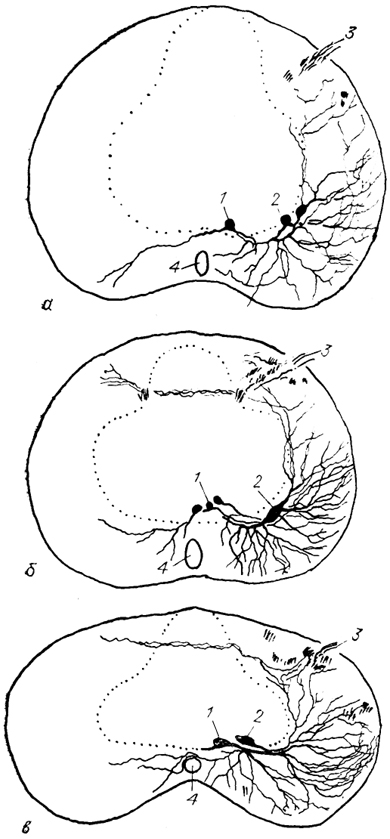
Рис. 21. Нейронная структура моторных ядер шейных сегментов спинного мозга хвостатых амфибий из группы саламандр: Rhyacotriton olympicus (a), Ptethodon jordani (б), Batrachoseps attenuatus (в) (Toth e. a., 1985).
1, 2 - медиальная (1) и латеральная (2) группы мотонейронов; 3 - дорсальный корешок; 4 - спинномозговой канал. Точками обозначена зона серого вещества.
клеток. Они образуют аксо-аксональные контакты электротонического или смешанного типа с маутнеровскими нейронами. Вторая популяция представлена мелкими вторичными мотонейронами, занимающими в мотонейронной колонне сегмента вентральное положение. Отдельные первичные мотонейроны иннервируют определенные участки миомеров, а разные популяции мотонейронов иннервируют в миомере различные типы мышц: первичные - белые, а вторичные - и белые и красные мышцы. Отдельное мышечное волокно иннервируется несколькими мотонейронами - полинейронная иннервация. Электрические органы, имеющиеся у некоторых рыб (в частности, у электрических скатов, сомиков) и развивающиеся из производных осевой мускулатуры, иннервируются специальной популяцией мотонейронов, расположенных в центральной зоне спинного мозга и связанных друг с другом многочисленными электротоническими контактами сомато - соматического и дендро-дендритного типов.
У амфибий, как и
у других низших позвоночных, мотонейроны разделяются на два основных класса: первичные (крупные) мотонейроны, дендриты которых распространяются на большие расстояния, захватывая несколько сегментов и переходя на противоположную сторону спинного мозга, и вторичные (мелкие) мотонейроны, аксоны которых не контактируют с маутнеровскими волокнами.
У хвостатых амфибий популяции мотонейронов формируют две группы, или колонны: вентромедиальную и вентролатеральную. Первая содержит в основном более мелкие грушевидные клетки, а вторая представлена крупными веретеновидными нейронами, причем степень развития той или иной мотонейронной колонны сильно варьирует даже у представителей одной труппы животных (рис. 21). Иннервация мышц у хвостатых амфибий полинейронная, и популяции мотонейронов содержат как первичные, так и вторичные мотонейроны.
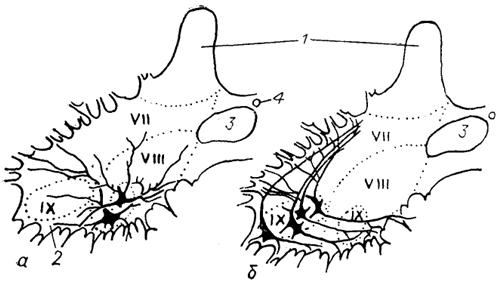
Рис. 22. Нейронная структура вентромедиального (а) и вентролатерального (б) мотонейронного ядра спинного мозга ящерицы (Cruce, 1979).
1, 2 -задние (1) и передние (2) рога спинного мозга; 3 - медиальные проприоспинальные тракты; 4 - спинномозговой канал; VII-IX -пластины Рекседа.
У бесхвостых амфибий в спинном мозге намечается пространственное разделение мотонейронных популяций, иннервирующих разные группы мышц. Клетки, иннервирующие осевую туловищную мускулатуру, расположены в мотонейронной колонне сегмента вентральнее и медиальнее мотонейронов, иннервирующих конечности, хотя внутри пула еще нет морфологической дифференцировки нейронов, иннервирующих отдельные мышцы. Мотонейроны образуют многочисленные электротонические, химические и смешанные контакты с клетками соседних сегментов и противоположной стороны спинного мозга, создавая основу для координированной деятельности мышц туловища и конечностей.
Мотонейроны спинного мозга рептилий организованы в две
моторные группы: вентромедкальную и вентролатеральную, нейроны которых отличаются друг от друга по размеру и характеру ветвления дендритов (рис. 22). Мотонейроны первой группы иннервируют осевую мускулатуру, второй - мускулатуру конечностей. У рептилий мотонейроны, иннервирующие отдельные мышцы, уже расположены раздельно. Однако в связи с тем, что у них самая сложная среди позвоночных мышечная система, имеется множество вариантов моторной иннервации. Так, красные медленные мышечные волокна рептилий иннервируются полинейронально и вдоль всей поверхности волокна, тогда как белые фазные волокна, разделяемые на быстрые и медленные, иннервируются мононейронально и локально. Очень часто в отдельных мышцах красные и белые волокна перемешаны друг с другом и иннервируются разными мотонейронами. Считается, что такая иннервация мышц у рептилий обеспечивает синхронизацию их работы (например, при движении туловища у змей). Подчеркнем, что у рептилий впервые среди позвоночных дендриты большинства мотонейронов не проникают на противоположную сторону спинного мозга, и это позволяет осуществлять более тонкую регуляцию деятельности мотонейронов спинномозгового сегмента через систему интернейронов.
Организация мотонейронных популяций, иннервирующих осевую мускулатуру млекопитающих, подробно изучена у крыс и кошек. Мотонейроны, связанные с определенными мышцами, занимают строго определенное положение в мотонейршном пуле. При этом мышцы, которые развиваются в соседних участках миомеров, иннервируются рядом расположенными нейронами, несмотря на возможное изменение положения данных мышц в туловище взрослого животного. У млекопитающих γ-мотонейроны, связанные с интрафузальными мышечными волокнами, рассеяны среди других нейронов данного пула. Интересно, что часть осевой мускулатуры млекопитающих - шейные мышцы - сохраняет филогенетически древний принцип иннервации - двумя разными типами мотонейронов, расположенных в разных ядрах. Остальная, как быстрая белая, так и медленная красная мускулатура не имеет двойной иннервации. Таким образом, в процессе эволюции сохраняются все возможные варианты организации нервно-мышечных отношений и соответственно мотонейронных пулов спинного мозга, причем часто независимо от филогенетического уровня.






