Наиболее характерная для среднего мозга всех позвоночных структура - зрительный тектум, tectum opticum, у анамний представляет собой самую заметную часть мозга (см. рис. 5). Строение тектума подробно изучено у большого количества видов, принадлежащих разным классам. Во всех случаях типичным оказывается четко выраженное ламинарное строение данного образования. Число слоев, нейронный состав и организация связей в значительной степени варьируют, что определяется не только принадлежностью животных к разным таксонам, но и уровнем развития систем мозга, в первую очередь зрительной сенсорной системы, и организацией моторных взаимодействий.
Уже в мозге круглоротых тектум является крупным образованием, выдающимся латерально и ограничивающим выстланный эпендимой тектальный желудочек. Несколько типов и нейронов
с хорошо развитым дендритным полем, ориентированным радиально или тангенциально, формируют 6-8 страт, разделенных на слои. Основной объем афферентов поступает в наружные отделы тектума и проводит информацию от органов зрения; в более глубоких отделах обнаружены входы от других сенсорных образований - октаво-латерального комплекса и спинномозгового ядра тройничного нерва. Имеются сведения и о входах от мозжечка и спинного мозга, таламуса и эпиталамуса.
Эфференты тектума образуют несколько путей: текто-бульбарный (связывающий его с ромбовидным мозгом), текто-таламичеекий и текто-гипоталамический. Часть аксонов тектальных клеток распределяется в пределах тегментума среднего мозга. Таким образом, уже в мозге круглоротых в строении тектума и организации его связей прослеживаются черты, общие для всех позвоночных. Существенным является и характер распределения проекций: верхние отделы являются основной мишенью для ретинальных входов, эфференты же формируются в глубоких отделах тектума.
В мозге пластиножаберных тектум образован несколькими слоями, составленными нейронами разных типов. В дорсовентральном направлении они подразделяются на четыре страты: str. medullare externum, cellulare externum, medullare internum, cellulare internum. Верхние отделы принимают зрительные афференты, в то время как к двум нижним стратам подходят проекции от других отделов мозга - ретикулярной формации, теленцефальных структур, а также от контрлатерального тектума. На одном виде показано и существование соматосенсорных проекций. Эфференты, возникающие в тектуме, формируют восходящие, комиссуральные и нисходящие связи: восходящие адресованы претектальной области и зрительным ядрам таламуса; комиссуральные связывают одноименные структуры и интерколликулярное ядро; нисходящие объединяются под названием текто-бульбарного тракта и адресованы в основном структурам ретикулярной формации и интерколликулярному ядру. Несколько противоречиво решается вопрос о существовании у пластиножаберных текто-спинального тракта, свойственного практически всем позвоночным: одни авторы отрицают наличие у акул прямых связей тектума со спинным мозгом, другие отмечают его чрезвычайно слабое развитие. Связи тектума и ядер глазодвигательного комплекса, по-видимому, опосредуются ретикулярными образованиями. В целом тектум пластиножаберных имеет значительные черты сходства с другими позвоночными, однако некоторые авторы в числе особенностей данной структуры у пластиножаберных отмечают менее выраженную стратификацию, относительную бедность клеточных форм и как примитивную черту билатеральный характер теленцефальных входов. Отличительной особенностью тектума является также
отсутствие ипсилатеральных проекций к ядру перешейка характерных для других позвоночных.
В мозге актиноптеригий тектум представляет собой наиболее значительную и высоко дифференцированную часть ЦНС. Степень его развития достаточно четко коррелирует с размерами глаз. Однако несмотря на то, что основной вход в эту структуру сформирован сетчаткой, вероятно, этим роль тектума не ограничивается. Об этом свидетельствует существование проекций от многих других отделов мозга. Кроме того, имеются виды, вторично лишенные зрительной системы, - пещерные слепые рыбы Astyanax hubbsi, у которых небольшой тектум оказывается сложно дифференцированным. Наиболее медиально расположенная часть тектума внедряется в полость желудочка и формирует возвышения, получившие название продольных валиков, torus longitudinalis. Эта структура, специфичная только для данного подкласса, варьирует от довольно крупного образования у Polyodon до небольшой клеточной группы в дорсомедиальной части тектума у Acipenser - среди хондростей, среди голостей она очень велика у Lepisosteus и чрезвычайно мала у Атiа у телеостей ее размеры не всегда коррелируют со степенью развития тектума.
В состав tectum opticum всех актиноптеригий входят шесть основных слоев (рис. 52): 1) str. marginale- наиболее развит у телеостей, наименее - у голостей (у хондростей он не описан); 2) str. opticum - содержит волокна оптического тракта и, по крайней мере у некоторых видов, аксоны другой природы (возможно, от тектума и претектальной области); 3) str. fibrosum et griseum superficiale - состоит из чередующихся слоев, составленных клеточными телами и их отростками, и является основным для ретинальных входов (кроме того, сюда подходят афференты от спинного мозга, ромбенцефалона и таламуса); 4) str. griseum centrale - эфферентный слой тектума, которому адресованы и комиссуральные проекции; 5) str. album centrale - через эту зону в тектум поступают афференты от спинного мозга, мозжечка и (у некоторых) теленцефалона; 6) str. griseum periventriculare - состоит из плотно лежащих нейронов с радиально ориентированными дендритами, пронизывающими весь тектум.
Основные афференты тектума актиноптеригий представлены волокнами зрительного тракта. Направляющиеся в тектум волокна, достигая хиазмы, полностью перекрещиваются.* Все ретинальные проекции топологически организованы, причем эта организация оказывается сходной у всех телеостей и подобна имеющейся у других позвоночных: представительство дорсовентрального направления сетчатки располагается в тектуме вентро-дорсального,
передняя половина сетчатки проецируется каудально, задняя - рострально. Другие входы в тектум образованы такими структурами, как спинной мозг, октаво-латеральная система, мозжечок, нисходящее ядро тройничного нерва, таламус, теленцефалон.
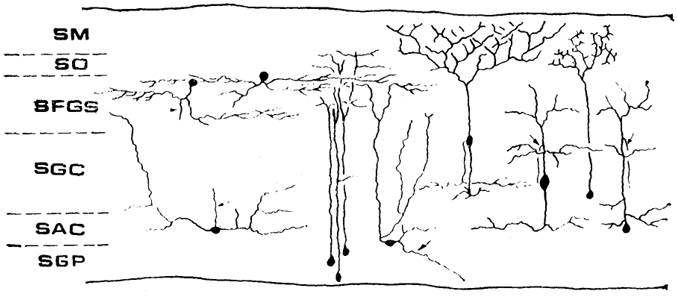
Рис. 52. Строение tectum opticum лучеперых рыб (Vanegas, 1980).
SM - stratum marginale, SO - stratum opticum, SFGS - stratum fibrosum et griseum super ficiale, SGC - stratum griseum centrale, SAC - stratum album centrale, SOP - stratum griseum periventriculare.
Эфференты тектума у актиноптеригий можно разделить на восходящие и нисходящие проекции. Восходящие направляются в претектальную область, латеральное коленчатое тело, дорсомедиальное оптическое и круглое таламические ядра. Помимо этих ипсилатеральных проекций обнаружены контрлатеральные входы к круглому ядру и небольшое число проекций к претектальной области и латеральному коленчатому ядру. Комиссуральные связи проходят через горизонтальную, поперечную и малую комиссуры (направление волокон, проходящих в составе задней комиссуры, остается невыясненным). Нисходящие волокна образуют два текто-бульбарных тракта - дорсальный и вентральный, trr. tectobulbaris dorsalis et ventralis. Дорсальный направлен в тегментум среднего мозга и перешеек (возможно, к его ядру), а также в полукруглый валик, torus semi-circularis, вентральный - в ретикулярную формацию ромбовидного мозга. Часть эфферентных проекций идет в мозжечок. Волокна, которые берут начало от вентролатеральной части тектума, вместе с другими церебеллопетальными связями от ростральных отделов (латеральное коленчатое тело, переднее таламическое ядро) образуют передний мезенцефало-церебеллярный тракт, tr. mesencephalocerebellaris anterior.
Тектум хвостатых амфибий наиболее прост и состоит из пе-ривентрикулярной зоны и поверхностного нейропиля. Последний образован ретинофугальными волокнами, а также входами от

Рис. 53. Строение тектума среднего мозга амфибий (Lazar, 1984).
1-9 - слои тектума; 10-16 - основные клеточные типы тектума: пирамидный (10), большой (11) и малый (12) пириформные, фузиформный (13), звездчатый (14), большой ганглионарный (15) амакриновый (16) нейроны.
других образований, в частности соматосенсорных. Самое поверхностное положение занимают проекции ядра перешейка. Афферентами, помимо названных, являются проекции от теленцефальных образований (латеральная амигдала, стриатум), некоторых ядер таламуса, претектальной области, ядра Даркшевича, а также от крупноклеточного ядра торуса, которое принимает информацию от первичных центров боковой линии. Эфференты тектума, составляющие восходящие проекции зрительного канала, адресованы дорсальному (ядро Беллончи и заднеталамическая область) и вентральному таламусу. Нисходящие проекции обеспечиваются волокнами tr. tectoisthmicus, в основном терминирующими на нейронах ядра перешейка, а также двумя текто-бульбарными трактами - прямым и перекрещенным, trr. tectobulbaris rectus et cruciatus. Первый оканчивается в непосредственной близости от мотонейронов продолговатого мозга, другой, вероятно, реализует воздействие через ретикулярную формацию.*
У бесхвостых амфибий строение тектума намного сложнее и в его пределах выделяют разное количество слоев, страт, зон (рис. 53). В типичном случае тектум имеет три зоны: поверхностную, центральную и перивентрикулярную. Последняя содержит тела клеток, в то время как остальные сформированы их отростками и приходящими афферентами. Характер связей и общий план строения тектума анура сходны с таковыми у
большой группы рептилий. В целом у амфибий с развитой зрительной системой в тектуме выделяют до 9 слоев. Входы от сетчатки преимущественно контрлатеральные (хотя недавно обнаружено и небольшое количество ипсилатеральных) и адресованы поверхностным слоям (6-9). В перивентрикулярную область приходят волокна от таламического зрительного центра, ядер дорсальных канатиков и ядер тегментальной области. Отметим, что число этих входов по сравнению со зрительными невелико. Эфференты через текто-таламический тракт направляются билатерально в латеральное коленчатое тело и заднелатеральную часть таламуса. Каудально идущие эфференты образуют текто-спинальный тракт, достигающий цервикального отдела спинного мозга. Наконец, как и у других животных, между тектумом и ядром перешейка имеются тесные двусторонние связи.
Тектум у рептилий изучен на большом количестве видов, и классификация его отделов проведена разными авторами по различным критериям. На основании эмбриологических данных, а также различия в связях тектум может быть подразделен на перивентрикулярную, содержащую около 80% клеточных элементов данной структуры, центральную и поверхностную зоны, zz. periventriculata, centralis et superficialis соответственно. Каждая зона представлена разным числом слоев (рис. 54). Общее число слоев (согласно одной из классификаций) - 14. Степень развития слоев и зон тектума, в первую очередь, связана с уровнем развития зрительной системы. Наиболее сложным оказывается tectum в мозге агам, игуан, хамелеонов, варанов - животных, отличающихся по своей экологии, но характеризующихся развитым зрением. Они имеют чрезвычайно развитый 14-й слой, занимающий верхнее положение в поверхностной зоне и составленный волокнами зрительного тракта.
Большинству рептилий свойствен, по-видимому, анцестральный тип развития тектума с хорошо представленными отделами и мало вариабельным наружным слоем (он характерен и для бесхвостых амфибий). Редукция перивентрикулярной зоны, обеднение ее клеточными элементами, слабое развитие наружного слоя - характерная особенность всех ныне живущих змей. Наконец, у видов с выраженной редукцией зрительной системы происходит потеря поверхностной зоны, а перивентрикулярная оказывается крайне бедна. Тем не менее и в этом случае тектум присутствует как самостоятельный отдел среднего мозга (рис. 55).
Основной объем афферентов тектума составляют аксоны ганглиозных клеток сетчатки. У рептилий это почти полностью контрлатеральные входы, поскольку перекрест в хиазме составляет около 90%. Для данного уровня характерна четкая ретинотопическая организация, причем с непропорционально большой
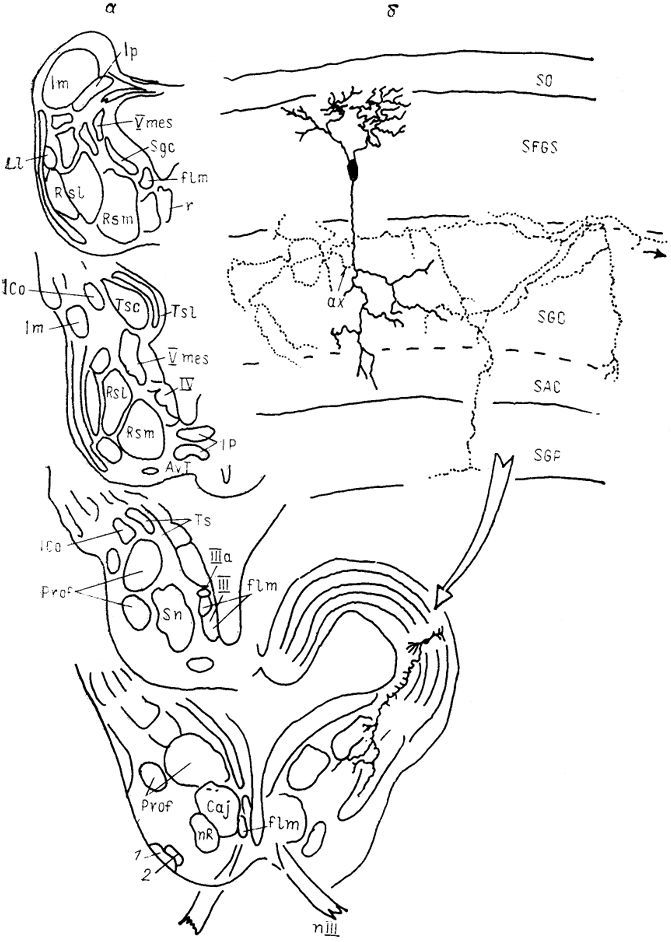
Рис. 54. Строение среднего мозга черепахи Pseudemys scripta (Sereno, 1985).
а - ядра среднего мозга, б - характер ветвлкния эфферентного нейрона тектума; 1, 2 - базальный оптичсекий тракт (1) и его ядро (2). Остальные обозначения, как на Рис. 52.
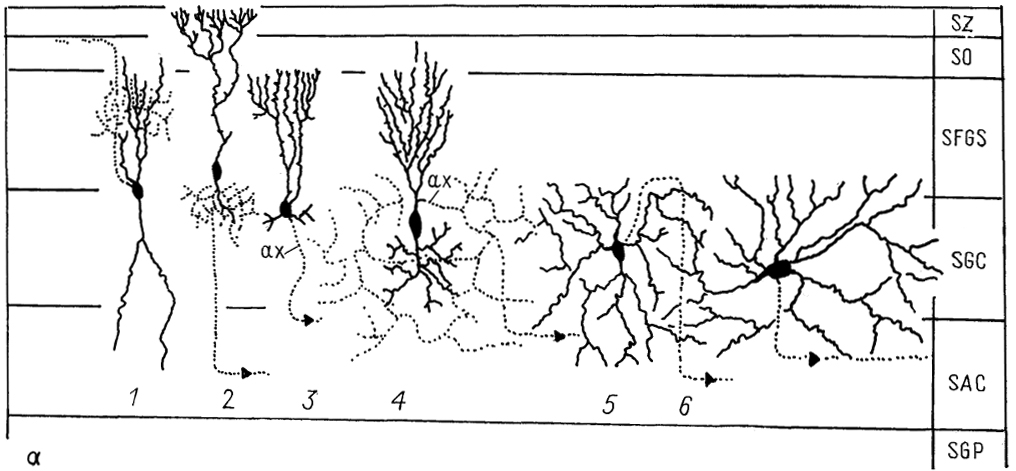
Рис. 55. Структура тектума среднего мозга змеи Thatnnophis sirtalis (Dacey, Ulinski, 1986).
a - эфферентные нейроны, б - интернейроны и афферентные волокна; 1-6 - эфферентные нейроны: текто-геникулятный (1), текто-истмальный (2), текто-истмо-бульбарный (3), текто-ротундальный (4), текто-бульбарные прямой (5) и перекрещенный (6); 7-10 - различные типы интернейронов; 11-16 - афферентные волокна из: сетчатки (11), претектума (12), вентральной части латерального коленчатого тела (13), вентро-латерального ядра таламуса (14) - перешейка (15) и глубокого ядра среднего мозга (16), SZ - stratum zonale. Остальные обозначения, как на рис. 52.

площадью представительства центральной части сетчатки: размеры центральной части сетчатки около 10°, а зоны ее представительства - 20%. Столь высокий магнификационный фактор обеспечивает разрешающую способность, на порядок превышающую таковую других областей. Помимо ретино-тектальных входов особенностью тектума являются интертектальные связи, степень выраженности которых зависит от бинокулярного перекрытия. Четко пространственно организованные связи характеризуют афференты ядра перешейка, что некоторыми авторами связывается с возможностью получения входов от ипсилатерального глаза. Кроме названных афферентами являются проекции вышележащих структур зрительной системы, в частности дорсальной коры и некоторых отделов таламуса. Кроме того, показаны входы от претектальной области и тегментального ядра среднего мозга. Роль последних, возможно, заключается в опосредовании влияний стриатума и во включении, таким образом, тектума в моторные системы мозга.
Существенной особенностью для понимания функциональной роли и эволюционных преобразований данного отдела является наличие ярко выраженной конвергенции разномодальных входов на нейронах тектума. Так, глубоким слоям тектума адресованы соматические входы, сохраняющие свойственную им в других отделах топическую организацию.* Полное контрлатеральное представительство позволяет предположить, что афференты образованы тригемино- и спинно-тектальными трактами. Судя по экспериментальным данным, акустическая информация проецируется на данный уровень от торуса, однако детальная пространственная организация этих входов неясна. В тех случаях, когда у рептилий обнаруживается инфракрасночувствительный ямковый (Якобсонов) орган, в контрлатеральный тектум приходят и распределяются в пределах 7-го слоя афференты либо непосредственно от специализированного ядра тройничного нерва (у питона), либо опосредованно - от ретикулярного ядра (nucl. reticularis caloris), получающего входы от первичного медуллярного центра (у гремучей змеи). У таких животных в 7-м слое тектума обнаруживается "карта пространства", воспринимаемого специализированными рецепторами. Отметим, что часть нейронов этого отдела может быть активирована как инфракрасным излучением, так и зрительными раздражителями.
Эфференты тектума рептилий формируют восходящие, нисходящие и интертектальные связи (при этом часть из них может обеспечиваться одними и теми же элементами). Первые представляют собой проекции к вышележащим отделам зрительной системы (в основном в дорсальный таламус), последние
обеспечивают бинокулярные взаимодействия и связь с претектальной областью. Нисходящие сформированы трактами, идущими к крупноклеточному ядру перешейка или его гомологу и образуют контакты с медуллярной ретикулярной формацией и спинальными нейронами по крайней мере цервикальных сегментов (рис, 54, б).
У птиц в пределах тектума насчитывают до 15 слоев (по одной из классификаций), первый слой - на поверхности. Особенностью является весьма редуцированный 15-й слой, расположенный в перивентрикулярной области и примыкающий непосредственно к эпендиме. Еще одна особенность, согласно некоторым авторам, заключается в существовании у птиц за пределами тектума двух групп клеток, которые по структуре связей рассматриваются как гомологи тектальных слоев других позвоночных. Одна из этих групп представляет собой интерколликулярное ядро, а вторая принадлежит ретикулярной формации.
Тектум птиц получает афференты от различных образований зрительной сенсорной системы: сетчатки, областей теленцефалона, связанных с обработкой зрительной информации, вентральной части латерального коленчатого тела, претектальной области, ядра перешейка. К другой группе образований, проецирующихся на тектум, относятся голубое пятно, тегментальное педункулярное ядро, ретикулярная формация и тегментальное педункуло-понтийное ядро.
Эфференты тектума птиц образуют группу восходящих и нисходящих проекций, а также адресованы ядру перешейка. Восходящие проекции распределяются среди таламических образований (круглое ядро, вентральная часть латерального коленчатого тела, задний отдел таламуса) и ядер претектальной области. Нисходящие связи направляются к моторным ядрам и ретикулярной формации в составе текто-бульбарного тракта.
Гомологом тектума у млекопитающих являются верхние (передние) холмы, colliculi superiores, - парные возвышения, занимающие ростральную часть поверхности среднего мозга. В их составе насчитывают 7 слоев (1-й расположен субпиально, 7-й граничит с центральным серым веществом среднего мозга). Среди клеточных форм много элементов, подобных имеющимся у рептилий, однако существуют и отличия. Нейроны 4-7-го слоев являются источниками текто-бульбарного тракта. На основании -сравнительного анализа характера развития и становления связей полагают, что нейроны этих слоев подобны клеткам слоев 3-5 перивентрикулярной области рептилий.
У млекопитающих лишь верхние отделы передних холмов являются специфическим образованием зрительной сенсорной системы: сюда подходят волокна зрительного тракта, проекции зрительной области коры и вентральной части латерального коленчатого тела. Таким образом, степень развития зрительной сенсорной системы оказывается решающей для выраженности
верхних отделов передних холмов млекопитающих. Эфференты указанной области адресованы небольшому числу зрительных центров: дорсальной части латерального коленчатого тела, nucl. parabigeminalis (вероятному гомологу ядра перешейка), а также комплексу ядер таламуса LP-Pul. Источниками этих связей являются клетки, сосредоточенные в нижних слоях поверхностной зоны.
Промежуточная и глубокая зоны передних холмов являются областью конвергенции входов зрительной, слуховой и соматической сенсорных систем. Заметим, что и для двух последних на уровне передних холмов отмечен упорядоченный характер проекций ("карты") рецепторных поверхностей. В указанные области приходят афференты и от других отделов: новой коры (преимущественно от соматосенсорных полей), черной субстанции, клиноподобного ядра ретикулярной формации, мозжечка. Эфференты данной зоны более многочисленны, чем поверхностной. У некоторых животных обнаружено до 40 структур, принимающих информацию от этого отдела. Основными эфферентными путями являются ипсилатеральные текто-понтийный в текто-бульбарный тракты и контрлатеральный текто-спинальный тракт. Часть путей адресована структурам, имеющим тесные связи с мозжечком: нижней оливе, дорсолатеральному мостовому ядру, ретикулярному тегментальному ядру моста. Другой мишенью колликулярных проекций служит комплекс глазодвигательных ядер, информация к которым проходит через претектальную область и ретикулярную формацию. Наконец, часть связей адресована таламическим ядрам, а через них полям 7 и 8 новой коры.
Таким образом, множественность проекций, особенности нейронного состава, отличия функциональных характеристик позволяют рассматривать глубокие слои передних холмов как самостоятельное образование, наиболее сходное с ретикулярной формацией среднего мозга. Анализ характера распределения связей показывает, что данный отдел играет важную роль в-обеспечении системных реакций (движений глаз, ушных раковин, головы - за счет доступа к моторным отделам) и, следовательно, является одним из элементов интегративных систем мозга млекопитающих.
В целом в строении тектума млекопитающих и птиц обнаруживается большое сходство как по типам составляющих их элементов, так и по соотношению связей с образованиями зрительной системы. В то же время если у млекопитающих передние холмы могут быть разделены, исходя из распределения связей, на поверхностные (зрительные) и глубокие (эфферентные) слои, то у птиц столь четкого деления провести не удается. Таким, образом, остается неясным вопрос о существовании у птиц гомолога тектальной области млекопитающих.
Передние холмы значительно варьируют по размерам и форме у разных млекопитающих: они малы у землеройки и чрезвычайно велики у утконоса, тупайи, кролика и др. В пределах одного отряда также возможны большие вариации, что, вероятно, объясняется не столько уровнем филогенетического развития, сколько другими факторами. Среди последних прежде всего - степень развития и совершенства зрительно-моторных взаимодействий, поскольку для передних холмов мозга млекопитающих характерны множественные связи, ориентированные преимущественно на исполнительные образования (комплекс глазодвигательных нервов, моторные ядра продолговатого мозга). Удельный вес проекций зрительного тракта у них снижается по сравнению с другими животными, поскольку сильно развит ретино-таламический вход. Кроме данного фактора весьма важна особенно у млекопитающих роль тех отделов, где взаимодействуют разные сенсорные системы и берут начало нисходящие тракты, адресованные не только ядрам черепномозговых нервов (текто-бульбарный тракт), но и спинному мозгу и ядрам моста (текто-спинальный и текто-понтийный тракты). Степень развития этий нисходящих систем не всегда зависит от совершенства зрительной системы. Так, у животных с крайне бедными верхними, получающими ретинальные входы слоями (например, летучие мыши) передние холмы могут достигать значительных размеров за счет высоко дифференцированных нижних слоев, напротив, у.некоторых видов с объемными передними холмами нисходящие тракты выражены незначительно. Возможны вариации в соотношении текто-спинальных и текто-понтийных волокон. Число последних, как правило, не превышает количества спинальных входов и весьма мало у животных с крупным мозгом (киты, гоминиды, слоны), у многих мелких животных, даже обладающих не очень развитым мостом, число этих волокон оказывается весьма значительным (летучие мыши, тупайи, кролики). Таким образом, степень развития передних холмов мозга млекопитающих, по-видимому, определяется целым комплексом факторов. Среди них весьма важен общий уровень развития интегративных систем мозга, в состав которых входит и область, традиционно называемая глубокими слоями передних холмов (по аналогии с другими позвоночными) и которую правильнее отделять от собственно зрительной части этого отдела и рассматривать как особую тектальную область. Подробное изучение связей этой зоны, проведенное на некоторых видах, позволило высказать предположение о ее принадлежности ретикулярной формации.
В целом тектум - самое характерное образование мозга позвоночных, представляющее собой ламинарную структуру, составленную чередующимися слоями клеточных тел и волокон. Для всех позвоночных основным сенсорным входом в эту область являются ретинальные волокна. У большинства животных они входят поверхностно и затем направляются вглубь, терминируя на отростках тектальных нейронов. У млекопитающих и пластиножаберных ситуация обратная: относительно глубоко входящие волокна распространяются в верхних слоях. В ходе эволюции возрастает как число клеток, так и число клеточных форм и меняется их распределение внутри структуры: если у низших основной объем центральной и поверхностной зон составлен волокнами, а клеточные тела сосредоточены в перивентрикулярной области, то у. высших наблюдается значительная миграция клеток в поверхностные отделы. Несмотря на сходство в общей организации гомологизация слоев тектума не проведена. Гомологичность же отдела в целом и его зон - перинентрикулярной, центральной, поверхностной - не вызывает сомнений.
На всем протяжении эволюции область тектума - это не. только центральный отдел зрительной системы позвоночных, но ж субстрат конвергенции входов от разных сенсорных систем. В этом смысле само название - tectum opticum - нельзя признать адекватным. Так, в тектуме разных позвоночных обнаруживаются "карты" сетчатки, улитки, внешних покровов, а в случае существования специализированных систем - и электрорецептивной зоны и рецепторов инфракрасного излучения. Таким образом, на данном уровне имеется представительство контрлатеральной части пространства, воспринимаемого разными типами рецепторов. Кроме того, для многих позвоночных обнаружены вестибулярные и проприоцептивные входы, однако их организация изучена менее подробно.

Рис. 56. Схема типичного ветвления одного волокна дорсального (а) и вентрального (б) текто-бульбарных трактов у черепахи Pseudemys scripta (Sereno, 1985).
1 - супрапедункулярное ядро, 2 - прерубральное поле.
Выраженные проекции к моторным отделам мозга обеспечивают тектуму решающую роль в организации таких реакций, как поворот в сторону добычи, предшествующий выбрасыванию языка у лягушки, определение местоположения теплокровной добычи гремучими: змеями, сочетанные движения глаз и головы, а: у млекопитающих, кроме того, - ушей и вибрисс.
Подобные реакции становятся возможными: благодаря множественности проекций к премоторным центрам, в первую очередь к ретикулярной формации, а также: к сегментарным моторным отделам - связи, которые носят название текто-бульбарных проекций. Для большинства позвоночных описаны дорсальный, промежуточный ш вентральный текто-бульбарные траокты, trr. tecto-bulbaris dorsalis, inter-medius, ventralis, соответственно.
Дорсальный текто-бульбарный тракт у всех позвоночных относительно постоянен (рис. 56, а). После выхода из тектума его волокна дают ипсилатеральную восходящую ветвь к вентральному
таламусу (у костистых рыб и высших позвоночных), затем основной пучок переходит на противоположную сторону, образуя предорсальный пучок, fasc. predorsalis (текто-спинальный тракт млекопитающих), который, располагаясь вентрально от медиального продольного пучка, спускается к медиальной ретикулярной формации и спинному мозгу, отдавая по пути многочисленные коллатерали. В ряде случаев показано, что источниками восходящих и нисходящих проекций являются одни и те же элементы. У млекопитающих текто-спинальный тракт значительно уступает в развитии другим нисходящим проекциям (рубро-, гипоталамо-, кортико-спинальным) и не превышает по степени развития таковой у рептилий. Наименее выражен он у приматов, наиболее - у хищных, а максимального для млекопитающих развития достигает у кошек и енотов - факт, не получивший исчерпывающего объяснения: такие очевидные факторы, как уровень развития зрительной системы и совершенство зрительно-моторных координации не коррелируют со степенью выраженности этого нисходящего пути у млекопитающих.
Промежуточный текто-бульбарный тракт содержит ипсилатеральный компонент, хорошо развитый у хрящевых рыб, рептилий и птиц. Его волокна адресованы глубокому ядру среднего мозга, латеральной части ретикулярной формации, ЦСВ среднего мозга. У млекопитающих большинство авторов не выделяет этот путь как самостоятельный, однако тектальные проекции к ретикулярной формации понтийного уровня и клиноподобному ядру среднего мозга (подобные обеспечиваемым промежуточным трактом других позвоночных) у них хорошо выражены.
Вентральный текто-бульбарный тракт (рис. 56, б) у позвоночных наиболее вариабелен. Он лишен восходящей ветви. У птиц и млекопитающих его волокна образуют значительное число терминалей на нейронах ядер моста, чем обеспечиваются текто-церебеллярные связи. Отметим, что у рептилий местом его окончания служит небольшое мелкоклеточное ретикулярное ядро, расположенное вблизи ядра перешейка и проецирующееся в свою очередь на мозжечок, - возможный прекурсор мостовых ядер.
Организация связей в пределах тектума оказывается весьма сложной. Для многих афферентов характерно закономерное распределение не только по слоям, но и в пределах слоя. Формируемые таким образом афферентные модули различаются по протяженности и ориентации. Интересно и то, что в некоторых случаях описано достаточно точное совпадение афферентных и эфферентных модулей. Так, например, у млекопитающих подобное совпадение обнаруживается для тригемино-тектальных и текто-тригеминальных связей. В то же время наиболее характерной является ситуация, когда тектальные эфферентные модули "захватывают" области нескольких афферентных, что типично и для текто-спинального тракта. Наконец, обнаружены области полного перекрытия двух разных модулей - гипоталамического афферентного и текто-оливарного эфферентного. Отметим, что в отношении текто-ретикулярных связей картина совершенно иная: для них характерно распределение терминалей по разным отделам и диффузная локализация источников этих
связей в пределах эфферентных слоев тектума. Таким образом, в отличие от других проекций, текто-ретикулярные не имеют топологической организации.
Традиционные представления о роли тектума заключались в рассмотрении его как высшего центра регуляции моторных ответов на телецептивные стимулы. Действительно, выраженность и сложная организация нисходящих связей этого образования с моторными отделами мозга, а также наличие разномодальных сенсорных входов позволяют уверенно говорить о принадлежности тектума к интегративным отделам мозга. При этом у низших позвоночных его роль оказывается решающей во многих отношениях. У высших же с дальнейшим развитием коры большого мозга увеличивается объем входов от теленцефальных образований, однако многие поведенческие реакции на сенсорные стимулы по-прежнему обеспечиваются именно активацией данного отдела мозга. Отметим, что наряду с возрастанием числа нисходящих связей от теленцефалона к тектуму у млекопитающих обнаруживается и значительное число восходящих проекций этого уровня к промежуточному и конечному мозгу, причем не только к структурам зрительной системы. Пока остаются неясными функциональное значение этих связей и их выраженность у других высших позвоночных.
Другим отделом, свойственным мозгу всех позвоночных, является полукруглый валик, torus semicircularis, или торус. У низших позвоночных эта структура находится между тектумом и покрышкой, а составлящие ее клетки лежат слоями в перивентрикулярной области (рис. 57). Положение торуса в мозге рептилий меняется: структура смещается каудально по отношению к тектуму, ростральные отделы торуса расходятся латерально и лишь каудальные полюса остаются вблизи средней линии. Дальнейшее развитие этого процесса приводит к формированию пластинки четверохолмия млекопитающих, где гомологичное торусу образование - задние холмы - располагается каудально от гомологов тектума - передних холмов, У птиц гомологичный отдел - дорсальная часть среднемозгового латерального ядра.
В мозге круглоротых и пластиножаберных торус представлен возвышениями на латеральной части вентрикулярной поверхности (у некоторых видов видимых возвышений не образуется). Его составляют мелкие гранулярные клетки субэпендимного слоя и лежащие в глубине более крупные нейроны. Уже у этих животных обнаруживаются общие для позвоночных черты организации связей торуса. Входы обычно сформированы октаво-латеральной областью продолговатого мозга, в зависимости от уровня развития которой варьирует и степень развития торуса. Ряд специфических черт в строении торуса обнаруживается в случае специализации октаво-латеральной области - у низших это прежде всего связано с электрорецепцией.

Рис. 57. Основные отделы среднего мозга хрящевых Scyliorhinus canicula (a), Raja clavata (б) и костистых Lepomis (в) рыб (Smeets, 1981, Wullimann, Northcutt, 1988).
Так, у скатов эфференты электрорецептивного дорсального и механорецептивного медиального ядер продолговатого мозга адресованы разным отделам (ядрам) торуса: латеральному, дорсомедиальному и вентромедиальному. Первый получает только проекции от дорсального ядра и связан с электрорецептивной системой, два других - от органов боковой линии и внутреннего уха. Вообще у пластиножаберных не найдено гомолога мезенцефалического акустического центра. Эксперименты с использованием дезоксиглюкозы позволили доказать, что вентромедиальное ядро торуса связано с обработкой акустической информации, однако его связи с центрами продолговатого мозга неясны. Дорсомедиальное ядро активируется под воздействием в конечном счете информации от механорецепторов органов боковой линии. Латеральное же ядро, принимающее информацию от электрочувствительного отдела продолговатого мозга, повидимому, нельзя рассматривать как гомологичное электрорецептивной зоне костистых рыб, поскольку у последних система электрорецепции, вероятно, не гомологична таковой других водных позвоночных.
У актиноптеригий полукруглый валик состоит из ряда слоев, параллельных вентрикулярной поверхности. Его размеры коррелируют с уровнем развития октаво-латеральной системы. При этом каждый из отделов торуса связан с обработкой информации, поступающей в конечном итоге от разных типов рецепторов октаво-латеральной системы. Так, у сома Ictalurus торус состоит из медиально расположенного центрального ядра и латерального ядра; последнее делится на части: латеральную и медиальную - меньшую по размерам. Акустический вход адресован центральному ядру, медиальная часть латерального ядра в основном содержит "электрочувствительные" нейроны, латеральная
же часть является местом проекций механорецепторов органов боковой линии. Еще более сложно дифференцирован торус у мормирид, у которых в его составе выделяют 7 ядер. Таким образом, входы медуллярных образований, относящихся к октаво-латеральной системе, на уровне торуса оказываются четко пространственно расположенными. При этом для каждого из отделов показана соматотопическая организация. Например, ростро-каудальное расположение рецепторов соответствует аналогичному направлению представительства в контрлатеральном торусе у сома Ictalurus. Эфференты торуса у актиноптеригий выяснены недостаточно. У голостей и телеостей они в составе заднего мезенцефало-церебеллярного тракта адресуются телу и; заслонке мозжечка. Комиссуральные связи образуют поперечную комиссуру, com. transversa, лежащую вентрально от промежуточного мозга. Полагают, что часть ее волокон адресована постхиазматической области гипоталамуса.
У бесхвостых амфибий торус представлен крупными парными утолщениями, выступающими в полость среднемозгового желудочка. Они очень велики у Ranidae (см. рис. 30) и смыкаются по средней линии, деля каудальную часть желудочка на дорсальный и вентральный отделы. В составе торуса у разных амфибий выделяют от трех до пяти ядер. У Rana catesbeiana торус включает главное, nucl. principalis, ламинарное, nucl. laminaris, крупноклеточное, nucl. magnocellularis, комиссуральное, nucl. commissuralis, субэпендимное, nucl. subependimalis, ядра. У Xenopus выделены только первые три. Афферентами ядер торуса являются входы от нижележащих образований слуховой системы. Так, главное ядро получает проекции от






