Все проводящие пути спинного мозга сосредоточены в белом веществе, разделяемом на три канатика: задний., боковой и передний. Основной объем канатиков занимают супраспи-нальные тракты, осуществляющие двустороннюю связь спинного
и головного мозга. Тракты, занимающие узкую полоску вокруг серого вещества, называются проприоспинальными (рис. 15).
Проприоспинальные тракты образованы аксонами интернейронов промежуточной зоны серого вещества. Распределение тел и окончаний проприоспинальных нейронов у позвоночных, начиная с самых ранних этапов эволюции, строго упорядочено. У высших позвоночных аксоны нейронов латеральных отделов пластин V-VII проходят в боковом канатике и оканчиваются в дорсолатеральных областях латеральных моторных ядер, ин-нервирующих мышцы-сгибатели. Нейроны центра пластины VII проецируются в вентромедкальную область этих ядер, связанную с мышцами-разгибателями. Интернейроны из медиальных районов пластин VII и VIII проецируются в медиальные моторные ядра, которые иннервируют осевую туловищную мускулатуру. При этом нейроны, образующие длинные восходящие и нисходящие проприоспинальные волокна, располагаются преимущественно в вентральных участках пластин V-VIII, а их аксоны оканчиваются в вентромедиальных районах пластин V-VII.
Проприоспинальные нейроны, относящиеся к разновидности ретикулярных изодендритных нейронов, создают морфологическую основу для широкой конвергенции на их телах и дендритах супраспинальных и сенсорных афферентов. Большинства проприоспинальных нейронов активируется супраспинальными трактами моносинаптически, а первичными афферентами - полисинаптически. Таким образом, появление и развитие в эволюции позвоночных системы проприоспинальных межсегментарных связей обеспечивает (наряду с другими механизмами) координацию сложных моторных реакций.
Супраспинальные тракты наибольшего развития и сложности достигают у высших млекопитающих - приматов, у которых описано их более двух десятков (рис. 15). Остановимся на наиболее важных из них.
Дорсальные канатики содержат восходящие тракты, волокна которых образованы аксонами сенсорных нейронов и коллатералями интернейронов пластин V и VI. Система дорсальных канатиков у высших позвоночных построена по соматотопическому принципу: чувствительные волокна от нижней части туловища и нижних конечностей проходят медиально, формируя тонкий пучок Голля (fasciculus gracilis), от верхней части тела и верхних конечностей - латерально, образуя клиновидный пучок Бурдаха (fasciculus cuneatus), причем в самих трактах пучки волокон от определенных участков тела и конечностей проходят также отдельно друг от друга. Функционально они тоже разделяются: волокна, несущие информацию от рецепторов волосяных фолликулов и конечностей, составляют поверхностную часть пучков, от мышечных рецепторов – среднюю
часть, а от рецепторов давления и вибрации - глубокую центральную часть. Оканчиваются волокна дорсальных канатиков в соответствующих ядрах продолговатого мозга.
В эволюции позвоночных происходит морфологическое обособление дорсальных канатиков от латеральных. Так, уже у хрящевых и некоторых костистых рыб они разделяются волокнами нисходящего бульбо-спинального тракта. Волокна дорсальных канатиков достигают продолговатого мозга и оканчиваются в зоне, которая по своей структурно-функциональной организации соответствует ядрам этого тракта у высших позвоночных. У наземных позвоночных уровень дифференцировки дорсальных канатиков тесно связан с развитием конечностей. Например, среди рептилий он наиболее, высок у крокодилов и наименее - у змей. Кроме того, начиная с амфибий, в организации этой системы все более отчетливо проявляется соматотопический принцип, Достигающий своего максимального выражения у млекопитающих и человека.
Система восходящих путей латеральных канатиков включает в себя ряд крупных трактов, соединяющих спинной мозг с продолговатым - спинно-ретикулярные, со средним - спинно-тектальные, с промежуточным мозгом - спинно-таламические, и мозжечком - спинно-мозжечковые. У низших позвоночных еще нет анатомического разделения трактов. У рептилий латеральные канатики уже дифференцированы на дорсо- и вентролатеральные пучки. Первый содержит в основном спинно-ретикулярные, спинно-тектальные и спинно-мозжечковые тракты, а второй - спинно-таламические и большую часть спинно-ретикулярных трактов.
Спинно-ретикулярные тракты обнаружены у всех изученных позвоночных - от круглоротых до человека. Они оканчиваются в каудальных отделах ретикулярной формации продолговатого мозга, моторных ядрах черепномозговых нервов, достигая в ряде случаев ретикулярной формации среднего мозга. Считается, что спинно-ретикулярные тракты позвоночных обеспечивают проведение информации от болевых рецепторов, а также пролриорецепторов.
Спинно-тектальные тракты описаны у большинства позвоночных, за исключением миног, лучеперых рыб и бесхвостых амфибий (возможно, что они утратили их в процессе эволюции независимо друг от друга). Спинно-тектальные тракты оканчиваются в центральном сером веществе мозга, в глубоких слоях тектума, тегментуме и интерколликулярном ядре.
Спинно-таламические тракты (спинно-таламический и спинно-ретикуло-таламический) обеспечивают болевую и температурную чувствительность и проведение информации от волосяных фолликулов, рецепторов давления и мышечных афферентов III группы. Спинно-ретикуло-таламичесюий тракт, как филогенетически наиболее древний, описан практически у всех
позвоночных, тогда как прямой спинно-таламический тракт обнаружен лишь у амниот и хрящевых рыб. Объем спинно-таламического тракта, а также локализация нейронов, образующих его, место окончания и количество передаваемой информации значительно варьируют даже у близкородственных групп позвоночных. Так, у млекопитающих проекции в специфические зоны таламуса (вентробазальный комплекс ядер) образованы нейронами пластин I и V спинного мозга, а в неспецифические зоны (медиальный комплекс ядер) -нейронами VII и VIII пластин. У птиц количество спинно-таламических нейронов очень невелико, полностью отсутствуют проекции из I пластины, при этом практически отсутствуют спинно-таламические проекции из шейных сегментов спинного мозга, иннервирующих крылья.
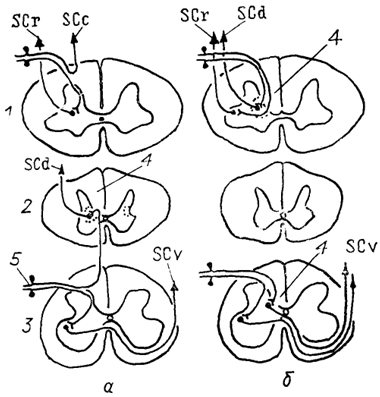
Рис. 23. Организация спинно-мозжечковых трактов у млекопитающих (а) и птиц (б) (Necker, 1989).
1-3 - шейный (1), грудной (2) и поясничный (3) отделы спинного мозга; 4 - ядро Кларка; 5 - чувствительные нейроны спинномозгового ганглия. Остальные обозначения, как на рис. 15.
Спинно-мозжечковые тракты есть у всех позвоночных, за исключением миног и миксин. У низших позвоночных спинно-мозжечковые волокна вместе с другими восходящими трактами входят в состав латерального канатика белого вещества спинного мозга и отделяются от них только в продолговатом мозге на уровне ядра V нерва. Разделение на вентральный и дорсальный компоненты намечается уже у хрящевых рыб и завершается у млекопитающих у которых выделяют дорсальный, вентральный и ростральный спинно-мозжечковые тракты и спинно-кунео-мозжечковый тракт (рис. 23). Дорсальный тракт начинается от крупных нейронов ядра Кларка в пластине IV серого вещества, оканчивается в I-IV и частично в V дольках мозжечка и несет информацию от отдельных мышц туловища и нижних конечностей. Вентральный образован нейронами нижних сегментов спинного мозга, расположенными в пластине V, и связан с проведением информации от группы мышц. Ростральный спинно-мозжечковый и спинно-кунео-мозжечковый тракты получают информацию от рецепторов верхних конечностей
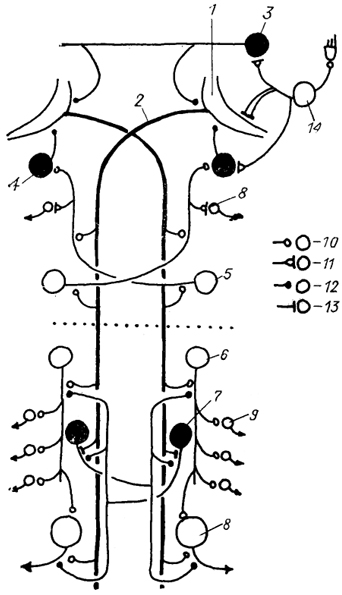
Рис. 24. Схема нейронной сети маутнеровских нейронов в спинном мозге костистых рыб (Faber e. а., 1989).
1 - маутнеровская клетка и ее аксон (2); 3-5 - комиссуральные (3),коллатеральные (4) и краниальные (5) интернейроны продолговатого мозга; 6, 7 - нисходящие (6) и реципрокные (7) интернейроны спинномозгового сегмента; 8, 9 - первичные (8) и вторичные (5) мотонейроны; 10-13 - синаптические контакты химического (10), электрохимического (11), химического тормозного (12) и электротонического (13) типов; 14 - нейроны чувствительного ганглия вестибулярного нерва. Темные клетки - тормозные, светлые - возбуждающие. Пунктиром обозначена граница головного и спинного мозга.
и начинаются в шейных сегментах спинного мозга. Все тракты и особенно дорсальный имеют четкую соматотопическую организацию. У птиц организация спинно-мозжечковых трактов иная, чем у рептилий и млекопитающих (рис. 23, б).
Нисходящие супраспинальные тракты опосредуют влияние головного мозга на моторные центры спинного мозга, и их активность связана прежде всего с такими рефлекторными актами, как плавание, бег, шагание, прыжки, полет. Система нисходящих трактов сформировалась на ранних этапах эволюции позвоночных и имеет единый план строения, включая несколько основных трактов: ретикуло-, вестибуло-, текто- и рубро-спинальные.
У низших позвоночных основную массу нисходящих трактов составляют ретикуло-спинальные тракты. Среди нейронов, образующих эти тракты, особо выделяются мюллеровские и маутнеровские клетки, расположенные в стволе мозга и обладающие длинными и нисходящими аксонами, которые моносинаптически связаны с мотонейронами спинного мозга, управляющими движением туловища и хвоста. У костистых рыб аксон каждой маутнеровской клетки ипсилатерально образует химические возбуждающие синапсы на спинальных нейронах двух типов: первичных мотонейронах и так называемых нисходящих интернейронах, аксоны которых на протяжении двух и более сегментов контактируют с первичными и вторичными мотонейронами (рис. 24).
Анализ структуры ретикуло-спинальных трактов высших позвоночных
показал, что данная система является наиболее консервативной. Это подтверждается ранним формированием ретикуло-спинальных трактов в онтогенезе.
Вестибуло-спинальные тракты развиты практически у всех позвоночных и разделяются на несколько пучков. У млекопитающих описаны прямой тракт (из латерального вестибулярного ядра) и два перекрещенных (из латерального и нисходящего вестибулярных ядер). Стимуляция этих ядер оказывает билатеральное действие на нейроны спинного мозга, связанные с движением головы, плечевого пояса и верхних конечностей. Тракт заканчивается на мотонейронах или интернейронах пластин VI-VIII.
Текто-спинальные тракты берут начало из различных отделов tectum opticum и опосредуют его влияние на мотонейроны спинного мозга. У млекопитающих они частично перекрещены. Обширные текто-спинальные тракты описаны у хрящевых и двоякодышащих рыб, амфибий (хвостатых и бесхвостых) и рептилий. По-видимому, они отсутствуют у некоторых костистых рыб (например, у лососевых). У круглоротых (по крайней мере, у миног) текто-спинальные волокна достигают верхних сегментов спинного мозга.
Перекрещенный рубро-спинальный тракт, сформированный нейронами красного ядра, обнаружен у бесхвостых и хвостатых амфибий, рептилий, птиц и млекопитающих; он, вероятно, отсутствует у безногих амфибий, змей и рыб. Прямой рубро-спинальный тракт описан у некоторых хрящевых (например, у скатов) и костных рыб, которые используют свои парные грудные придатки для передвижения. Таким образом, рубро-спинальные тракты эволюционно связаны с появлением конечностей и поясов конечностей, и степень их развития коррелирует со степенью развития мозжечка (однако присутствие рубро-спинального тракта у змеи Nerodia fasciata опровергает мнение о том, что он может быть связан только с регуляцией движений конечностей). Заканчиваются тракты в спинном мозге на интернейронах пластин V-VII.
Кортико-спинальный (пирамидный) тракт появляется у птиц и млекопитающих и является наиболее молодой из всех проводящих систем спинного мозга. Размер тракта увеличивается в ряду млекопитающих, тесно коррелируя с развитием моторной активности вида и со способностью к тонким дифференцированным движениям конечностей, и достигает максимума у человека. В спинном мозге зона прямых проекций тракта значительно варьирует у разных видов (рис. 25). У низших млекопитающих (например, у сумчатых) кортико-спинальные волокна достигают только шейных и грудных сегментов, проходя в дорсальных канатиках, и оканчиваются в задних рогах спинного мозга. У высших млекопитающих кортико-спинальный

Рис. 25. Окончание кортико-спинального тракта в шейных сегментах спинного мозга опоссума (а), кошки (б), макаки (в), шимпанзе (г) и характер ветвления одного волокна в моторных ядрах мозга макаки (д) (Shinoda е. а., 1979; Kuypers, 1981).
1, 2 - задние (1) и передние (2) рога спинного мозга; 3, 4 - латеральный (3) и передний (4) кортико-апинальные тракты; 5 - моторные ядра.
тракт проходит вдоль всего спинного мозга, разделяясь на два! компонента (латеральный и вентральный).
Кортико-спинальные волокна начинаются в разных областях новой коры, и, таким образом, пирамидный тракт по существу представляет собой комплекс отдельных путей, каждый из которых отличается по характеру окончаний в спинном мозге. Так, у высших приматов волокна от моторных зон коры проецируются в промежуточную зону и моторные ядра, волокна от соматосенсорных зон - в пластины I-IV. Дифференцированное распределение кортико-спинальных проекций свидетельствует об их влиянии на разные спинальные механизмы, вплоть до регуляции сенсорных афферентов в пластинах I и II. Более того, волокна из моторных зон коры оканчиваются преимущественно на телах и проксимальных дендритах мотонейронов, тогда как проекции из соматосенсорных полей - на дистальных дендритах. У большинства млекопитающих пирамидный тракт созревает в постнатальный период, причем в процессе его развития происходит элиминация многих кортико-спинальных
волокон, выходящих из некоторых корковых зон (например, зрительной коры).
Таким образом, уже в строении спинного мозга различных позвоночных отчетливо прослеживаются тенденции последовательной дифференциации его элементов и связей, а также усложнение межнейронных взаимодействий, обусловленные системным повышением уровня организации ЦНС.
ГЛАВА 3
ПРОДОЛГОВАТЫЙ МОЗГ
Продолговатый мозг, Medulla oblongata, уже у круглоротых представлен в достаточно дифференцированном виде. Являясь продолжением спинного мозга, он на значительном протяжении сохраняет свойственные последнему структурные особенности и форму. Изменение конфигурации и взаимного расположения структур продолговатого мозга у всех позвоночных происходит в дорсальных отделах в результате формирования IV желудочка мозга путем расщепления крыловидной пластинки и расхождения ее в вентролатеральных направлениях. Рострально желудочек переходит в полость среднего мозга. Верхняя его часть сформирована хориоидной крышей и у большинства позвоночных на всем протяжении ограничена мозжечком.
ОБЩИЙ ПЛАН СТРОЕНИЯ ПРОДОЛГОВАТОГО МОЗГА
Основной объем ядер продолговатого мозга составляют центральные проекции V-XII пар черепномозговых нервов. В зависимости от уровня филогенетической или адаптивной эволюции выраженность этих отделов варьирует, что и обусловливает значительное расхождение во внешнем виде и топографии медуллярного отдела. При анализе строения продолговатого мозга традиционно рассматривают локализацию ядер по нескольким колоннам, параллельным оси мозга. Границы их образованы бороздами на стенках желудочка, дополняющими пограничную борозду в период развития мозга и сохраняющимися в ряде случаев у взрослых животных. Эти борозды разделяют серое вещество на зоны, которые являются продолжением соответствующих участков спинного мозга (рис. 26).
Соматическая афферентная зона представлена в продолговатом мозге соматосенсорной (система тройничного нерва и
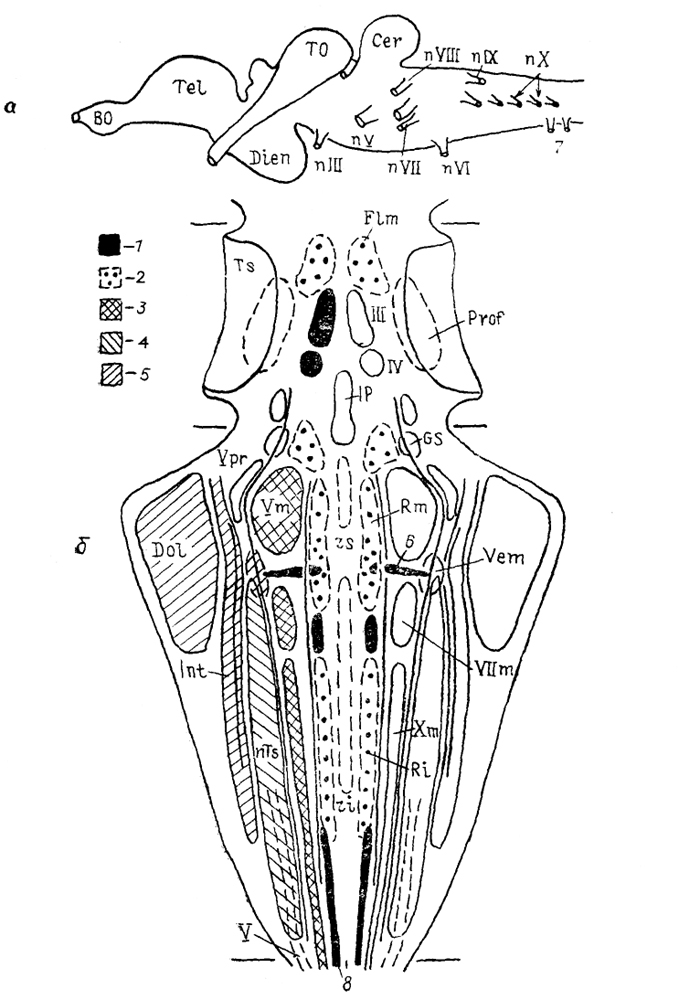
Рис. 26. Строение (а) и расположение (б) важнейших ядер стволовой части головного мозга анамний (Nieuwenhuys, Meek, 1985).
1, 3-5, 8 - колонны: соматомоторная (1), висцеромоторная (3), висцеросенсорная (4), специальная соматосенсорная (5), моторная спинного мозга (8); 2 - ретикулярная формация; 6 - маутнеровская клетка; 7 - спинно-окципитальные нервы.
ядра дорсальных канатиков) и специализированной октаво-латеральной (система вестибуло-кохлеарного нерва и в ряде случаев - водные позвоночные - часть лицевого нерва) областями. Висцеросенсорная зона содержит ядра языкоглоточного, лицевого и блуждающего нервов, связанные с обработкой информации от вкусовых и висцерохимических рецепторов.
Соматомоторная зона объединяет многочисленные клетки, составляющие на уровне продолговатого мозга ядра отводящего и подъязычного нервов. Эта колонна продолжается и на уровне среднего мозга, где она содержит ядра глазодвигательного и блокового нервов. Каудально она переходит в моторную колонну спинного мозга. Висцеромоторная зона образована ядрами тройничного, лицевого, языкоглоточного и блуждающего нервов.
Внутренняя топография того или иного отдела и его компонентов чрезвычайно вариабельна, и далеко не всегда структурная организация продолговатого мозга четко соответствует вышеизложенной. Так, анатомические границы зон часто не совпадают с границами колонн в силу различного развития структурных элементов, их образующих. Кроме того, во многих группах животных отмечается гипертрофия одних отделов и слабое развитие других. В результате специализация сенсорных систем, отражаясь в строении продолговатого мозга, приводит к значительным изменениям его конфигурации, причем у низших позвоночных эти вариации заметнее, чем у высших.
Вместе с тем наличие ярко выраженной специализации какой-либо системы оказывается не только интересной иллюстрацией адаптивных возможностей мозга, но и позволяет вычленить (при сравнительном анализе) базовые признаки, характеризующие те или иные отделы мозга. В этом отношении одной из наиболее интересных групп является надкласс Рыбы, где в зависимости от условий сред обитания и соответствующей специализации сенсорных и моторных систем обнаруживается чрезвычайно широкий диапазон вариантов строения разных отделов мозга и в первую очередь медуллярного уровня (рис. 27). В подобных случаях наблюдаются разные модификации внутреннего строения - от простого увеличения числа элементов до чрезвычайно сложных клеточных дифференциаций и четко выраженного ламинарного характера строения стволовых образований (рис. 28).
Естественно, что в состав продолговатого мозга входят и структуры, не относящиеся к сегментарным отделам, но включенные в различные системы мозга. Их удельный вес на уровне продолговатого мозга невелик, однако в ходе филогенеза происходит его неизбежное возрастание. Наконец, значительная часть продолговатого мозга составлена ретикулярной формацией (см. гл. 6).
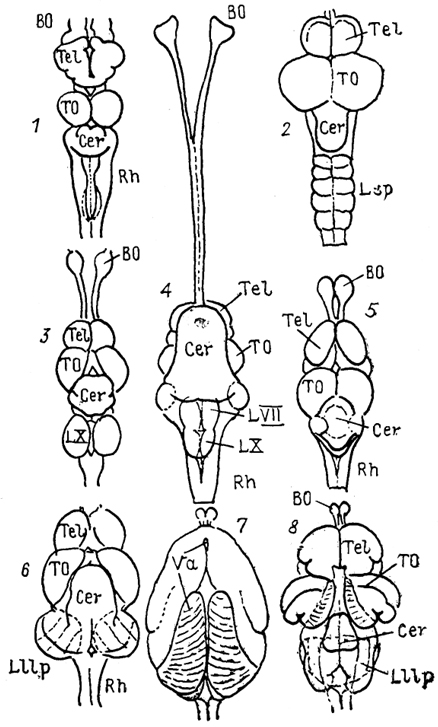
Рис. 27. Общий вид мозга разных представителей лучеперых рыб (Nieuwenhuys, Meek, 1985).
l - Amia, 2 - Trigla, 3 - Carassius, 4 - Clarlas, 5 - Xenomystus, 6 - Eigenmannia 7, 8 - Gnathonemus: до (7) и после (8) удаления заслонки.
Система черепномозговых нервов развивается как продолжение системы спинномозговых нервов, осуществляющих иннервацию туловища соответственно расположению миотомов.
Черепномозговые нервы в свою очередь формируются как бранхиальные, осуществляющие иннервацию области жаберных щелей, и эволюционируют параллельно с эволюцией жаберных дуг.

Рис. 28. Строение ромбовидного мозга разных групп рыб (Nieuwenhuys, Meek, 1985).
a - расположение колонн в мозге; 6-е - фронтальные сечения мозга Amia (б), Clarlas (в), Carassius (г), Xenomystus (д), Eigenmannla (e). 1-3 зоны продолговатого мозга; специальная соматосенсорная (1), висцеросенсорная (2), висцеромоторная (3); 4, 5 - электросенсорная область представительства ампуллярных (4) и бугорковых (5) рецепторов органов боковой линии электрических рыб.
В целом у ранних позвоночных четко выделяются бранхиальные нервы, гомологичные будущим тройничному (V), лицевому (VII), языкоглоточному (IX) и блуждающему (X). Начинающийся впоследствии процесс усложнения этой системы приводит к ее значительным преобразованиям. Так, путем слияния проекций чувствительных ветвей VII, IX и X пар черепномозговых нервов дифференцируется система вестибуло-кохлеарного (VIII) нерва, представленного уже в мозге круглоротых (рис. 29). Вероятно, позднее, судя по его слабой выраженности у бесчелюстных, формируется центральное представительство исключительно моторного отводящего (VI) нерва (рис. 30). У рептилий в качестве черепномозгового выделяется подъязычный (XII) нерв, представляющий собой объединение передних корешков трех ростральных сегментов спинного мозга. Отметим, что у некоторых рыб этот нерв уже присутствует в виде так Называемой задней доли, lobus posterior, но расположен не в краниальном, а в спинальном отделе (см, рис. 5, 6, 7).
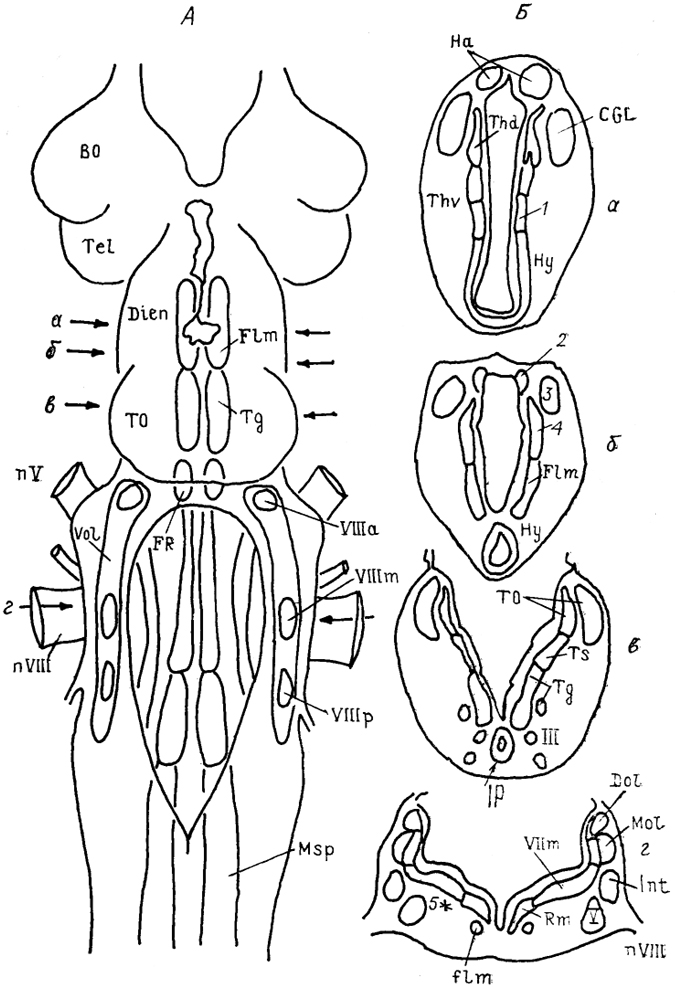
Рис. 29. Общий вид (А) и фронтальные сечения (Б) мозга миноги Ichthyomyzon unicuspis (Ronan, 1989).
1 - ядро заднего бугорка, 2 - субкомиссуральный орган, 3 - претекталъное ядро, 4 - ядро задней комиссуры, 5 - маутнеровская клетка; а-г - срезы в рострокаудальном направлении.
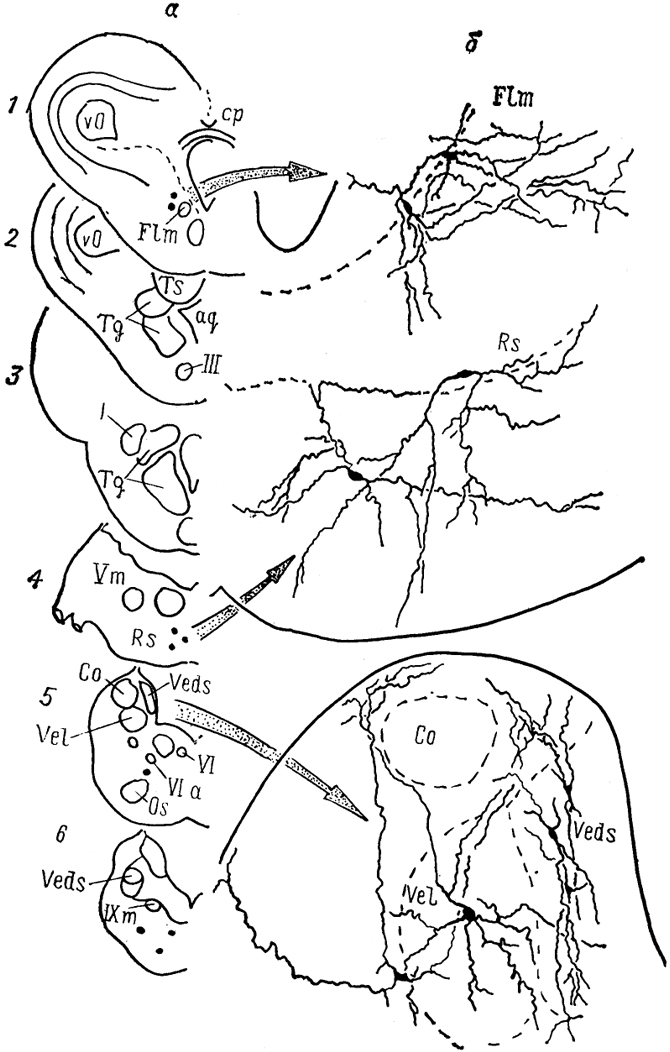
Рис, 30. Ядра (а) и нейронный состав (б) мозга лягушки Rana esculenta (Toth e. a., 1985).
1-6 - срезы в рострокаудальном направлении.
У бесхвостых амфибий местом его выхода является уровень II-III шейных сегментов. У рептилий, птиц и млекопитающих происходит вычленение добавочного (XI), чисто моторного нерва, как полагают, отделяющегося от блуждающего нерва.
Таким образом, в восходящем ряду позвоночных происходят процессы, приводящие к выделению и развитию имеющейся в настоящее время у высших системы черепномозговых нервов. Адаптивные изменения, обусловленные различием сред обитания, сказываются на всех компонентах этой системы, выражаясь не только в разной локализации выхода корешков из мозга, но и в разной степени развития их центральных проекций, а следовательно, в размерах (диаметре) нервов. На уровне продолговатого мозга такие различия особенно заметны в области производных крыловидной пластинки, где сосредоточены сенсорные ядра черепномозговых нервов.






