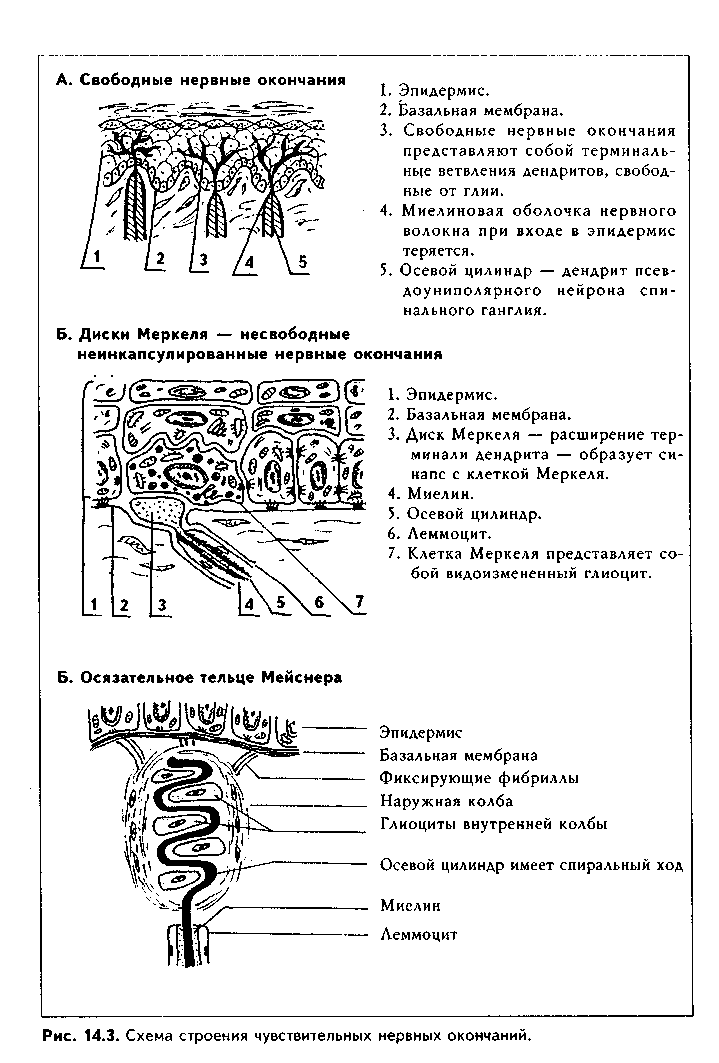
Представляют собой терминальные разветвления дендритов нейроцита. КЛАССИФИКАЦИЯ. Существует несколько принципов классификации рецепторных нервных окончаний.
1. По месту восприятия раздражителя. Рецепторные нервные окончания делятся на три группы: экстерорецепторы, воспринимающие раздражение из внешней среды; интерорецепторы, служащие для восприятия раздражений из внутренней среды организма; проприорецепторы, воспринимающие информацию от опорно-двигательного аппарата.
2. В зависимости от специфичности раздражения, воспринимаемого рецептором. Выделяют: механорецепторы, воспринимающие механические раздражители, перемещения частей тела; хеморецепторы воспринимают химические раздражители; терморецепторы улавливают изменения температуры, а ноцирецепторы воспринимают чувство боли.
3. По способу восприятия раздражителя выделяют контактные рецепторы, приходящие в состояние возбуждения при непосредственном воздействии на участок тела, и дистантные рецепторы, воспринимающие раздражитель, удаленный от организма (рецепторные клетки сетчатки глаза, органа слуха, обоняния).
4. Морфологическая классификация. В зависимости от строения все рецепторы делят на свободные и несвободные. Свободные рецепторные нервные окончания состоят только из конечных ветвлений дендрита чувствительного нейрона, а несвободные нервные окончания кроме терминалей нервного отростка имеют также и клетки олигодендроглии (леммоци-ты), которые окружают терминали дендрита и участвуют в восприятии раздражения. В свою очередь, несвободные нервные окончания делятся на неинкапсулированные (не окруженные по периферии соединительнотканной капсулой) и инкапсулированные, имеющие такую капсулу. Свободные нервные окончания воспринимают в основном болевые раздражения. Большинство несвободных нервных окончаний являются механорецепто-рами. В последнее время, однако, выдвигается небеспочвенная точка зрения, что не существует разделения рецепторов в зависимости от вида воспринимаемого раздражения, все рецепторы способны воспринимать раздражители любой модальности, а характер ощущения зависит от силы раздражителя.
МОРФОЛОГИЯ РЕЦЕПТОРОВ. 1. Свободные нервные окончания. Внаибольшем количестве представлены в коже. Это механорецепторы на волосяных фолликулах, ноцицептивные (воспринимающие болевые раздражители) нервные окончания в эпидермисе (рис. 14.3). Их много также в многослойном плоском неороговевающем эпителии, серозной оболочке. В эпидермисе они представлены древовидными ветвлениями дендритов псевдоуниполярных нейронов спинальпых ганглиев.
2. Несвободные неинкапсулированные нервные окончания представлены осязательными дисками Меркеля, а также нервными окончаниями соединительной ткани. Особенно много их в дерме. Осязательные диски Меркеля (рис. 14.3 б) состоят из отростка нервной клетки, который закан- чивается расширением в виде П. диска. Этот диск образует синапс с клеткой Меркеля, которая лежит в эпидермисе. В цитоплазме клетки Меркеля есть секреторные гранулы с нейромедиатором. Механическое раздражение вызывает выделение гранул из клеток Меркеля, их содержимое ведет к деполяризации отростка нейроцита.
Несвободные неинкапсули-рованные окончания в соединительной ткани построены следующим образом. Осевой цилиндр освобождается от миелина и на значительном расстоянии окружается глиальными клетками, тесно с ними контактируя. Очень часто на поперечном разрезе видна билатеральная симметрия таких окончаний.
3. Несвободные инкапсулированные нервные окончания построены по общему принципу. К этим окончаниям относятся нервные окончания в соединительной и мышечных тканях. Есть следующие разновидности этих окончаний: пластинчатые тельца Фатер-Пачини, осязательные тельца Мейснера, концевые колбы Краузе, генитальные тельца Догеля, тельца Руффини, нервно-мышечные и нервно-сухожильные веретена и др.

|
Наиболее распространены пластинчатые тельца Фатер-Пачини. Они встречаются в коже, молочной железе, в брыжейке, во внутреннил органах, около кровеносных сосудов, около суставов. Это крупные образования диаметром от 1 до 5 мм (рис. 14.4, 14.5). Имеют овальную форму и состоят из соединительнотканной капсулы, терминалей дендрита псевдоуниполярного нейрона и нейролеммоцитов (олигодендроглии). Дендрит при подходе ккапсуле теряет миелиновую оболочку и со всех сторон окружается нейролем-моцитами. Они формируют так называемую внутреннюю колбу. Эта колба снаружи покрыта соединительнотканной капсулой, которая часто называется наружной колбой. Капсула состой! из послойно параллельно лежащих кол-лагеновых волокон (образуют от 10 до 60 слоев) и клеток фиброцитов. В наружной капсуле встречаются кровеносные сосуды. Между наружной и внутренней колбами лежат специализированные отростчатые олигодендроглиоци-ты, контактирующие с осевым цилиндром. При давлении на тельце механическое воздействие во много раз усиливается слоями наружной колбы, что делает этот рецептор очень чувствительным. Давление смещает наружную колбу по отношению к внутренней. При этом раздражаются отростчатые олигоденд-роциты, передающие возбуждение на дендрит.

|
В сосочковом слое дермы обнаруживаются осязательные тельца Мейс-нера. Они являются механорецепторами и по размеру меньше телец Фа-тер-Пачини (50—140 мкм). Имеют овальную форму (рис. 14.3 и 14.6). Снаружи находится очень тонкая слоистая капсула — наружная колба. Дендрит псевдоуниполярного нейрона теряет миелиновую оболочку, разветвляется, и его ветви входят внутрь капсулы по спирали. Перпендикулярно к ним лежат глиальные клетки, которые вместе с терминалами дендритов образуют внутреннюю колбу. Незначительная деформация капсулы передается глиоцитам, которые имеют синаптическую связь с дендритом.
Концевые колбы Краузе являются барорецепторами и терморецепторами. Они лежат в дерме кожи, слизистых оболочках. Имеют небольшие (40—150 мкм) размеры. Также состоят из наружной капсулы и внутренней колбы. Внутренняя колба образована плоскими глиоцитами, между которыми проходят, формируя своеобразный клубочек, тонкие ветви дендрита. Наружная капсула очень тонкая.
Генитальные тельца Догеля находятся в особо чувствительных областях кожи, в первую очередь, в области наружных половых органов, коже молочных желез. Они похожи по строению на колбы Краузе, но в в отличие от них в тельце входят несколько отростков от нескольких нейро-цитов. Поэтому раздражение генитального тельца вызывает сильную иррадиацию возбуждения.
Тельца Руффини находятся в соединительной ткани кожи и в капсулах суставов. Воспринимают чувство давления. Имеют вид верстеновид-ных образований длиной до 2 мм. Осевой цилиндр во внутренней колбе разветвляется с образованием большого количества ветвей с булавовидными утолщениями на конце. Капсула хорошо выражена.
В гладкой мышечной ткани чувствительные нервные окончания также инкапсулированы, они контактируют с группой гладких миоцитов.
В скелетной мышечной ткани чувствительные нервные окончания называются нервно-мышечными веретенами. Представляют собой инкапсулированные нервные окончания (рис. 14.7, 14.8). Наружная соединительнотканная капсула нервно-мышечного веретена окружает несколько тонких так называемых интрафузальных мышечных волокон. В отличие от обычных мышечных волокон, лежащих снаружи и называемых экстрафузаль-ными, интрафузальные волокна тонкие, содержат мало миофибрилл и имеют светлую цитоплазму. Различают два вида интрафузальных мышечных волокон (рис. 14.8). 1. ЯС-волокна. Ядра этих волокон лежат в центральной части мышечного волокна, образуя скопление в виде ядерной сумки (сокращенно ЯС). В месте расположения ядер волокно резко расширяется.
2. ЯЦ-волокна. Эти волокна имеют равномерную толщину, а ядра лежат но всей длине волокна в его центре, формируя ядерную цепь.
Вокруг данных двух видов интрафузальных волокон в их центральной части образуются специфические синапсы дендритов чувствительных нейронов в виде:
1) аннулоспиральных (кольцеспиральных) окончаний, в которых отростки нервных клеток закручены вокруг центральной части интрафузалыю-го волокна по спирали и на большом протяжении вступают в синаптичес-кую связь с ним; аннулосниральные окончания имеются как на ЯС-, так и на ЯЦ-волокнах. 2) гроздьевидных окончаний, которые находятся только на ЯЦ-волокнах. При этом они формируются не в центральной части, а на периферии волокна.

|
На интрафузальных волокнах имеются также двигательные нервные окончания, которые представлены аксонами у-мотонейронов передних рогов спинного мозга. Они регулируют длину интрафузальных волокон и поддерживают их тонус. Все свободное пространство между мышечными волокнами заполнено жидкостью и ограничено тонкой капсулой. Всякое изменение тонуса мышцы ведет к изменению давления жидкости в полости капсулы. При этом давление передается на дендриты. Аннулосниральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, а гроздьевидные — только на изменение длины. Благодаря нервно-мышечным веретенам организм по-
стоянно получает информацию о степени сокращения мышц, что формирует представление о положении тела в пространстве.
МЕЖНЕЙРОННЫЕ СИНАПСЫ
Это особый вид нервных окончаний, когда разветвления отростков одних нервных клеток заканчиваются на других нервных клетках. При помощи синапсов возбуждение передается с одной нервной клетки на другую.
Классификация синапсов. Существует несколько подходов к классификации синапсов.
i. По механизму передачи нервного импульса. Синапсы делятся на химические, электрические и смешанные. В химических синапсах возбуждение передается при помощи химического вещества — нейро-медиатора. Эти синапсы являются наиболее распространенными в нервной системе высших животных. В электрических синапсах потенциал действия передается прямо с мембраны одного нейрона на другой. Смешанные синапсы представляют собой сочетание признаков и химического, и электрического синапсов.
2. Морфологическая классификация синапсов. Учитывает особенности контактирующих участков иейроцитов. Различают аксо-соматичес-кие, аксо-дендритические, аксо-аксональные, дендро-дендричес-кие, сомато-соматические синапсы.
3. Физиологическая классификация. По вызываемому эффекту на нервную клетку синапсы делятся на возбуждающие и тормозные.
4. Медиаторная классификация синапсов. По химическому типу медиатора есть синапсы холинергические, аминергические (адренергические, серотонинергические, дофаминергические); пуринергичес-кие, аминокислотные (медиаторами являются аминокислоты: ГАМК, глицин, глутамат, аспартат и т.д.), пептидергические (см. ме-диаторную классификацию нейроцитов).
СТРОЕНИЕ СИНАПСОВ. Любой синапс состоит из трех частей: пре-синаптического полюса с пресинаптической мембраной, синапти-ческой щели и постсинаптического полюса с постсинаптической мембраной.

|
Электрические синапсы. Эти синапсы построены по типу нексусов: две мембраны (пре- и постсинаптическая) соседних нейронов тесно сближаются друг с другом до расстояния в 2 нм, и это место контакта пронизано многочисленными коннексонами. Следовательно, синаптическая щель в электрическом синапсе практически отстутствует. Коннексоны представляют собой своеобразную пору через обе мембраны, которая по краям ограничена особыми белковыми молекулами коннексинами. Коннексоны пропускают не только ионы щелочных металлов, играющих важную роль в формировании электрических потенциалов, но и молекулы с ММ 1000—2000. Поэтому кроме электрического сопряжения коннексоны дают возможность нейронам обмениваться метаболитами. В отличие от химических синапсов, в которых проведение сигнала несколько задерживается, в электрических синапсах импульс проводится практически без задержки и в обе стороны. Значение электрических синапсов неизвестно. Предполагают, что оно связано с необходимостью быстрого сопряжения нервных клеток.
Химические синапсы. В отличие от электрических, химические синапсы передают нервные импульсы только в одном направлении и с задержкой (синаптическая задержка). Это наиболее распространенный у млекопитающих тин синапсов. Химические синапсы имеют все три отчетливо выраженные составные компоненты: нресинаптический и ностсинаптический полюсы и синаптичес-кую щель (рис 14.9) (в световом микроскопе синапсы видны в виде пугов-чатых утолщений на нейроцитах, см. рис. 13.1).
В пресинаптическом полюсе находятся пресинаптические пузырьки с медиатором, митохондрии, агранулярная ЭПС, нейротрубочки и нейрофи-ламенты. Синаптические пузырьки имеют различное строение в зависимости от содержащегося в них медиатора. Так, пузырьки с ацетилхолином имеют мелкие размеры и электронно прозрачные. Синаптические пузырьки с но-радреналином крупнее и имеют в центре электронноплотную часть. Содержащие пептиды пузырьки имеют крупные размеры, плотную сердцевину и окружены периферическим светлым ободком.