

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Основы обеспечения единства измерений: Обеспечение единства измерений - деятельность метрологических служб, направленная на достижение...
Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов...
Интересное:
Искусственное повышение поверхности территории: Варианты искусственного повышения поверхности территории необходимо выбирать на основе анализа следующих характеристик защищаемой территории...
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Лечение прогрессирующих форм рака: Одним из наиболее важных достижений экспериментальной химиотерапии опухолей, начатой в 60-х и реализованной в 70-х годах, является...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
РВНСТ является самым распространенным видом соединительных тканей. Она сопровождает самые мелкие кровеносные сосуды, образуя строму паренхиматозных и входя в состав оболочек слоистых органов. Функциями РВНСТ являются: защитная, опорная, трофическая, регулятор-ная, гомеостатическая, формообразующая (участие в образовании формы органов), пластическая (участие в восполнении объема разрушенной части органов и тканей, в том числе при воспалении, регенерации и других защитных реакциях).
Строение. Как и все ткани мезенхимного происхождения, РВНСТ состоит из клеток и межклеточного вещества.
|
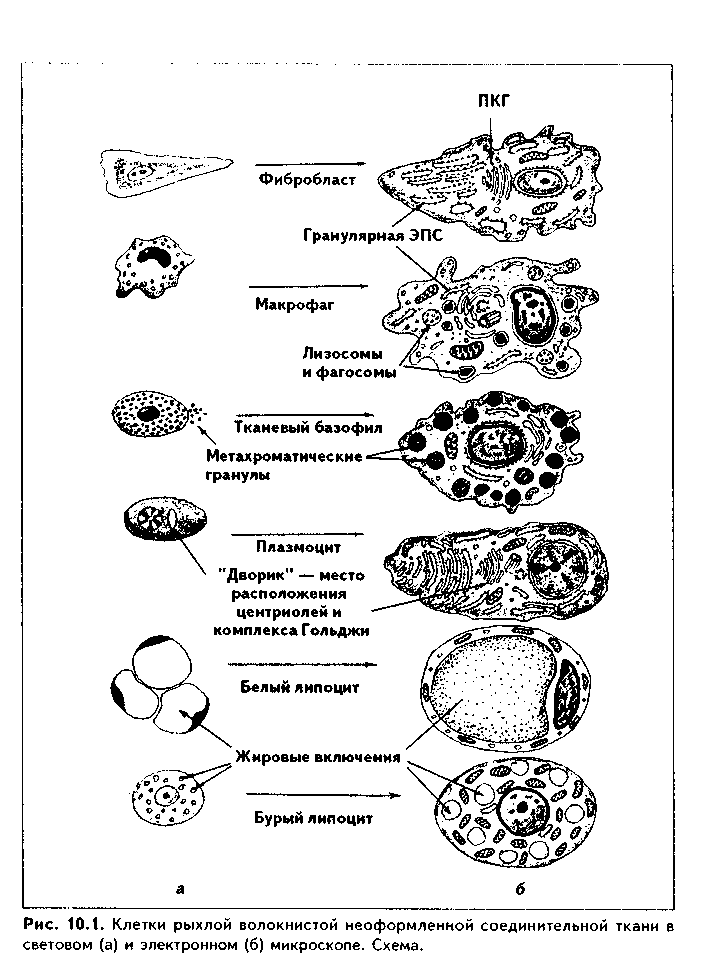
Клетки РВНСТ. Клеточный состав РВНСТ разнообразный и объединен в несколько дифферонов клеток (рис. 10.1).
1. Дифферон фибробластов (стволовые клетки -> полустволовые клетки —> малодифференцированные фибробласты —> дифференцированные фибробласты —> фиброциты; фиброкласты; миофибробласты).
2. Дифферон макрофагов (стволовая клетка крови... —> моноциты крови —> макрофаги РВНСТ).
3. Дифферон плазмоцитов (В-лимфоциты крови —> В-лимфоциты РВНСТ —> плазмобласты —> ироплазмоциты —> плазмоциты).
4. Дифферон тканевых базофилов (стволовая клетка крови... —» тканевой базофил РВНСТ).
5. Дифферон липоцитов (малодифференцированные фибробласты —> лииоциты).
6. Дифферон пигментоцитов (клетки-предшественницы —> пигмеи-тоциты).
7. Адвснтициальные клетки.
8. Перициты.
9. Лейкоциты.
Все клетки рыхлой соединительной ткани можно объединить в две группы:
1. Местные, или "собственные", клетки. Эта группа включает все вышеуказанные диффероиы клеток кроме лейкоцитов. 2. "Пришлые" клетки, или тканевые лейкоциты.
По источникам развития все клетки РВНСТ разделяются на 3 группы:
1. Клетки, относящиеся к линии механоцитов. К ним относятся
адвентициальные клетки (перициты), клетки дифферопа фибробластов и жировые клетки (липоциты). Эти клетки развиваются из особой стволовой клетки механоцитов костномозгового происхождения, отличной от стволовой кроветворной клетки.
2. Клетки, развивающиеся из стволовой кроветворной клетки макрофаги, плазмоциты, тучные клетки (тканевые базофилы), лейкоциты Оба вида стволовых клеток имеют мезенхимное происхождение, подвергаясь дивергентной детерминации.
3. Клетки нейроэктодермального происхождения развиваются из ганг-лиозных пластинок. К ним относятся пигментоциты.
ФИБРОБЛАСТЫ (ФБ). В эмбриогенезе фибробласты возникают непосредственно из мезенхимных клеток. В постнатальном онтогенезе источником их развития является стволовая клетка механоцитов соединительной ткани, которая находится в костном мозге и, поступая в соединительную ткань, превращается в более близкого предшественника — адвентициаль-ную клетку. Фибробласты — наиболее многочисленная популяция клеток РВНСТ. Их функция — образование межклеточного вещества РВНСТ — волокон и компонентов основного вещества.
При формировании дифферона фибробластов в процессе деления и дифференцировки адвентициальных клеток последовательно образуются:
1. Малодифференцированные (юные) ФБ — клетки с высоким ЯЦО, умеренным числом органелл, крупным или овальным базофильным ядром с 1—2 ядрышками. Эти клетки уже обладают определенной способностью к синтезу (коллаген, компоненты основного вещества), но отличаются также способностью к митозу. Следовательно, их функции — участие в биосинтезе межклеточного вещества и увеличение популяции ФБ.
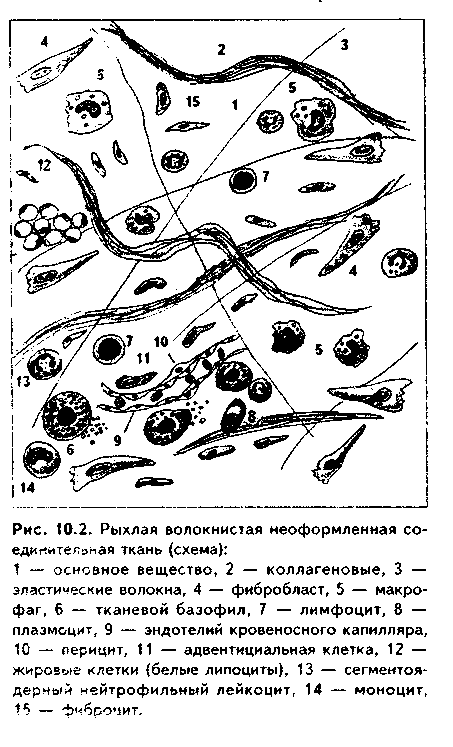
2. Дифференцированные (специализированные, или зрелые) ФБ имеют низкое ЯЦО, светлое, с эухроматином, ядро, в котором находятся 1—2 ядрышка (рис. 10.2). В световом микроскопе цитоплазма этих клеток слабо базофильна и подразделяется на внутреннюю, вокругъядерную, более ба-зофильную эндоплазму и наружную, более светлую эктоплазму. В эндоплазме находятся хорошо развитые органеллы белкового синтеза: гранулярная ЭПС, комплекс Гольджи, митохондрии, лизосомы (рис. 10.3). На периферии клетки, в эктоплазме, находятся развитые компоненты цитоске-лета, обеспечивающие формирование отростков и активное передвижение ФБ. Эктоплазма ФБ но восприятию красителей часто может не отличаться от основного вещества, как бы переходит в него без видимых границ.
Функциями специализированных ФБ являются: а) активный синтез компонентов межклеточного вещества; б) регуляторное влияние на функции других клеток соединительной ткани путем секреции большого количества медиаторов; в) разрушение коллагена, синтезированного самими фибробластами. Осуществляется как внутриклеточно, так и внеклеточно при помощи вырабатываемого фибробластами фермента коллагеназы. Разрушение синтезированного коллагена является необходимым процессом контроля состояния межклеточного вещества, поддержания тканевого гомео-стаза. Разрушая и синтезируя вновь коллаген, фибробласты также модифицируют, перестраивают рубцовую ткань.
|
3. Фиброциты. При старении фибробласты превращаются в фиброциты — неактивные клетки веретеновидной формы с плотным гипербазо-фильным ядром, узким ободком бедной органеллами цитоплазмы и сниженным белковым синтезом.
Функции. Фиброциты являются конечным этапом развития фибробла-стов и заканчивают свое существование путем апоптоза. Однако в последнее время показано, что фиброциты в определенной степени участвуют в обновлении компонентов межклеточного вещества, т.е в поддержании тканевого гомеостаза, а при травмах могут трансформироваться в клетки, приобретающие черты активных зрелых фиб-робластов.

|
4. Миофибробласты. Это ФБ, которые имеют сильно развитые сократительные филаменты и похожи на гладкие мио-циты, но в отличие от них не окружены базаль-ной мембраной и содержат более сильно развитую ЭПС и комплекс Гольджи. Светом и кроско-пически миофибробласты не отличаются от обычных гладких миоцитов.
Функции. 1. Эти клетки в большом количестве появляются при регенерации тканей, при этом сокращаются и сближают края раны (вызывают ее контракцию). Одновременно эти клетки активно синтезируют компоненты межклеточного вещества. Следовательно, за счет деятельности миофибробластов происходит более быстрое заживление ран. 2. Могут превращаться и гладкие миоциты и тем самым участвуют в репаративной регенерации гладкой мышечной ткани. В рубцовой ткани превращаются вначале в фибробласты, а затем в фиброциты. 5. Фиброкласты —
это ФБ, в которых сильно развиты лизосомы. В функциональном отношении похожи на макрофаги. Их функция — разрушение межклеточного вещества при его избыточном увеличении, например, в матке после родов, в рубцах после регенерации. Поскольку синтезирующие фибробласты также способны к фиб-роклазии, некоторые авторы рассматривают фиброкласты как дифференцированные фибробласты, у которых функция разрушения межклеточного вещества преобладает над синтетическими функциями. Таким образом, ФБ и фиброкласты являются функциональными антагонистами, регулирующими объем межклеточного вещества и гомеостаз РВНСТ.
Макрофаги (МФ). Это второй по численности после фиброблаетов дифферон РВНСТ. Развиваются из потомков стволовой кроветворной клетки — моноцитов крови после попадания их в ткани. Преобразование моноцитов в макрофаги сопровождается увеличением размеров клетки до 25—50 мкм, приобретением ядром бобовидной формы, накоплением лизосом и других орга-нелл: митохондрий, лизосом, эндоплазматичес-кой сети.

Гипертрофируется комплекс Гольджи, нарастает количество пиноцитозных пузырьков. Резко усложняется поверхность макрофага за счет увеличения количества микроворсинок, ямок и складок. Усиливается подвижность макрофагов, способность их к пино-цитозу и фагоцитозу, бактерицидная активность. В клетках нарастает активность лизосо-мальных ферментов, а также ферментов энергетического обмена. В соединительной ткани макрофаги могут находиться как в покоящемся (покоящиеся гистиоциты), так и в акшициты;, так и в активном состоянии (блуждающие гистиоциты). Морфологически эти две формы клеток существенно отличаются.В световом микроскопе макрофаги имеют бобовидное ядро и резко очерченные контуры цитоплазмы, в которой обнаруживаются вакуоли (рис. 10.1, 10.2). Покоящиеся гистиоциты обычно трудно отличить от фиброцитов, т.к. они имеют уплощенную форму, небольшие плотные ядро и цитоплазму, в которой содержится ограниченное количество органелл. Неактивные макрофаги обычно прикреплены к коллагеновым волокнам. Блу-жающие гистиоциты, напротив, высоко подвижны, что определяет высокую динамичность формы клеток. Поверхность их неровная, с многочисленными нсевдоподиями. При электронной микроскопии в блуждающих макрофагах выявляются множество лизосом, митохондрий, гладкая и гранулярная ЭПС, включения гликогена, фагоцитированные частицы (фаго-лизосомы) (рис. 10.4). На поверхности цитолеммы макрофаги несут многочисленные рецепторы для медиаторов иммунной системы, нейромедиаторов, гормонов и др., а также молекулы клеточной адгезии, позволяющие им взаимодействовать с другими клетками и межклеточным веществом, совершать миграционные процессы.

|
Функции. 1. Фагоцитарная функция: распознавание, поглощение и расщепление с помощью ферментов микроорганизмов и других антигенов, погибших клеток, старых компонентов межклеточного вещества тканей и др. 2. Антигенпредставляющая (презентирующая) функция: переработка антигена, перевод его в высокоиммунную форму и передача лимфоцитам. Благодаря этой функции макрофаги запускают иммунные реакции. В настоящее время антигенпрезентирующие клетки выделены в отдельную группу макрофагов, по ряду показателей отличающихся от блуждающих макрофагов, осуществляющих неиммунный фагоцитоз. 3. Секреция: медиаторов — веществ, регулирующих функции других клеток РВНСТ и им-мунокомпетентных клеток; противовирусных (интерферон) и противо-бактериальных (лизоцим, активные метаболиты кислорода и др.) факторов. 4. Участие в противоопухолевом иммунитете. 5. Регуляция тканевого гомеостаза. Макрофаги элиминируют старые элементы тканей, участвуют в тканевом обмене веществ, особенно в обмене жиров, регулируют состояние межклеточного вещества и активность тканевых клеток. 6. Регуляция регенерации: секретируют ряд веществ, стимулирующих заживление ран, участвуют в макрофагической фазе воспаления (см. ниже).
Тканевые базофилы (ТБ) (синонимы — тучные клетки, лаброциты, ма- стоциты). Третий по численности клеточный дифферон РВНСТ. Источником развития ТБ является стволовая клетка крови. ТБ образуются из одного предшественника с базофильными лейкоцитами крови, имеют с ними весьма схожие строение и функции, но не абсолютно идентичны. Полагают, что популяция ТБ в РВНСТ может пополняться за счет деления молодых тучных клеток.
В РВНСТ ТБ часто лежат возле кровеносных сосудов и нервов (рис. 10.1. 10.2).
|
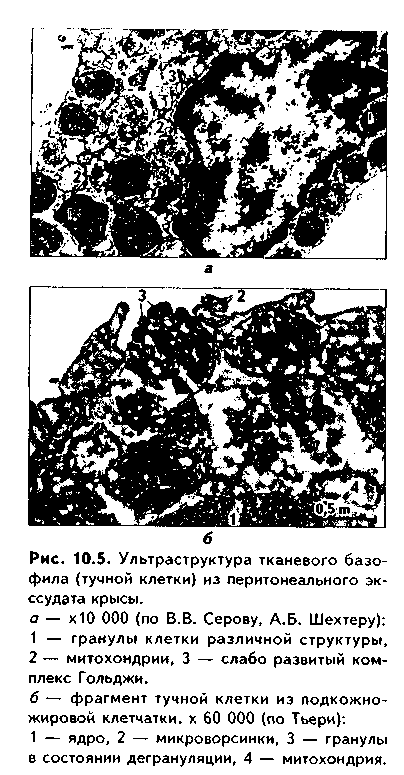
Имеют размеры от 10 до 30 мкм. Форма может быть различна: овальная, веретеновидная, неправильная и др. Ядра клеток округлые, с преобладанием гетерохроматина, маскируются гранулами и плохо различимы в световом микроскопе. В цитоплазме содержатся умеренно развитые органеллы общего назначения и компоненты цитоскелета. Встречаются также липидные включения. Характерная особенность — наличие большого количества метахроматических гранул (окрашивающихся в цвет, отличающийся от цвета красителя в растворе) (рис. 10.5). Гранулы тучных клеток отличаются от аналогичных базофилов крови не только большим количеством, но также большей вариабельностью формы и ультраструкту-ры: есть гранулы с плотным го могеиным и зернистым строением, а также кристаллоподоб-ныс. Мстахромазия гранул обусловлена гепарином, который снижает свертываемость крови, понижает проницаемость сосудов. Гранулы содержат также гнетами н (а у грызунов и серотонип). Эти вещества могут изменять состояние основного вещества РВНСТ, увеличивать проницаемость микрососудов. Кроме того, в состав гранул входят некоторые ферменты, хемотаксические факторы для эозинофилов и нейтро-филов и ряд других веществ. Секреция гранул тучными клетками называется дегрануляцией. Она может быть как медленной и незначительной по объему (в условиях нормы), так и быстрой, массивной (при аллергических и анафилактических реакциях).
Функции. 1. Регуляция тканевого гомеостаза (гомеоста-тическая) — проницаемости сосудов, свертываемости крови, трофики тканей. Осуществляется за счет медленной секреции содержимого гранул. 2. Синтез основного вещества РВНСТ (гепарина, хонд-роитинсульфатов, гиалуроповой кислоты, гликопротеииов). 3. Рсгуляториая функция. Заключается в регуляции функций других клеток РВНСТ и крови, а также состояния межклеточного вещества путем выделения медиаторов. 4. Участие в иммунных реакциях. Медиаторы тучных клеток регулируют функции клеток иммунной системы, силу иммунного ответа. Эти клетки осуществляют фагоцитоз комплекса антиген—антитело, поглощение избытка гистами-на. Они участвуют в аллергических и анафилактических реакциях. 4. Стимуляция регенерации тканей и участие в гисто- и органогенезе. В частности, тучные клетки стимулируют развитие волос. Плазмоциты (плазматические клетки). Разминаются из В-лимфоцитов кропи через такие стадии: В-лимфоцит -> илазмобласт -> проплазмоцит -> плазмоцит. При этом в клетке постепенно снижается ЯЦО, в цитоплазме накапливаются органеллы белкового синтеза. Вместе с В-лимфоцитами плазмоциты всегда в том или ином количестве содержатся в РВНСТ. Особенно большое их количество в РВНСТ собственных пластинок слизистых и серозных оболочек внутренних органов.

Плазмоциты имеют размеры 7-10 мкм (встречаются и более крупные плазмоциты размером до 20 мкм) и овальную форму с эксцентрично лежащим крупным овальным или округлым ядром (рис. 10.1). Хроматин в ядре создает картину колеса со спицами. Цитоплазма клеток сильно базо-фильна, однако около ядра имеется светлая неокрашенная часть («дворик») — место расположения комплекса Гольджи. При электронной микроскопии в цитоплазме сильно развита гранулярная ЭПС (рис. 10.6). Ее цистерны сильно уплощены и располагаются параллельно и достаточно
тесно друг к другу, развиты комплекс Гольджи, митохондрии. Плазмоциты секретируют без оформления секрета в секреторные гранулы. Однако иногда на светомикроско-пическом уровне в плаз-моцитах обнаруживаются тельца Русселя — плотные сферические включения, содержащие углеводы и белки, в том числе и иммуноглобулины. В электронном микроскопе тельца Русселя представляют собой гомогенный материал, лежащий в резко расширенных зонах гранулярной ЭПС. Полагают, что эти образования появляются при нарушении процессов сип-теза и секреции иммуноглобулинов.
Функции. Единственной функцией плаз- моцитов является выработка иммуноглобулинов - антител, инактивирую-щих антигены. Благодаря этой функции они участвуют в гуморальном иммунитете. Плазмоциты — единственные в организме клетки, синтезирующие антитела.
Жировые клетки (липоциты, адипоциты). Встречаются практически повсеместно, однако количество их в различных участках даже в одном органе может сильно варьировать от единичных до мощных скоплений.
Источником развития липоцитов являются малодифференцированныс фибробласты, в которых постепенно накапливаются лииидные включения, сливающиеся в одну жировую кайлю. Механизм отбора фибробластов, направляющихся по пути превращения в липоцит, неизвестен. Показано, что при голодании после истощения липидных включений жировые клетки могут вновь превращаться в фибробласты.
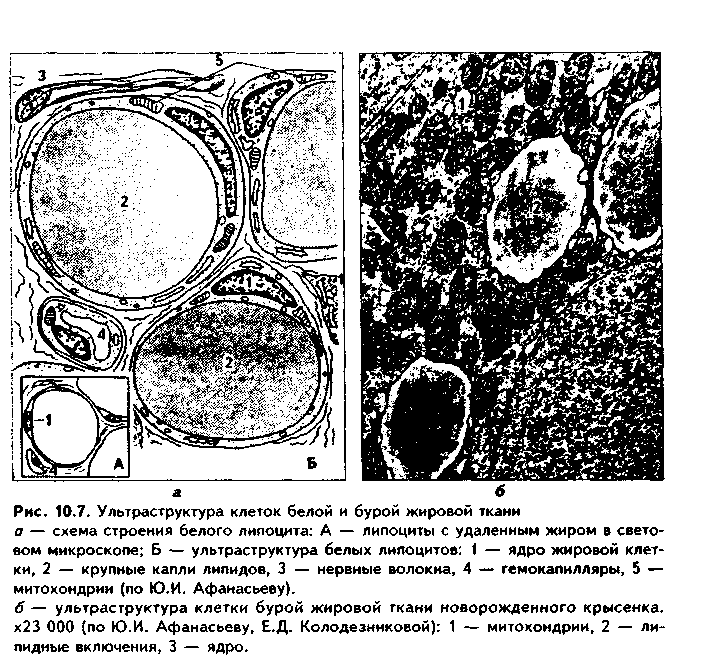
Различают белые и бурые липоциты (см. рис. 10.2, 10.7). Белые липоциты лежат группами возле гемокапилляров. Имеют перстневидную форму и крупные размеры. Ядро темноокрашенное, лежит на периферии. Цитоплазма в виде узкого ободка. В центре клетки — большая жировая капля (жировое включение), окрашивающаяся Суданом. При электронной микроскопии в цитоплазме выявляется мало органелл (рис. 10.7). Функциями белых липоцитов являются депонирование жира, воды (при распаде жира образуется ее большое количество), трофическая и терморегулирующая функции.
Бурые липоциты имеют меньшие размеры и многоугольную форму. Ядро расположено в центре, округлое. В цитоплазме содержатся множественные жировые капли. При электронной микроскопии в клетках имеются умеренно развитые ЭПС и комплекс Гольджи (рис. 10.7). Многочисленные митохондрии имеют сильно развитые кристы и сосредоточены вокруг липидных капель. Находящиеся в них железосодержащие окислительные ферменты цитохромы придают клеткам бурый цвет.
Главной функцией бурых липоцитов является функция выработки большого количества тепла (функция термогенеза), поскольку окисление-жиров в них сопровождается не синтезом АТФ, а выделением большого количества тепла. Это достигается за счет белка термогенина. разобщающего окисление и фосфорилирование. Второстепенной функцией является депонирование жиров.
Пигментоциты (пигментные клетки). Все пигментные клетки образуются из нейромезенхимы — нервного гребня. Содержат большое количество пигментных включений (включения меланина). Меланин обладает повышенной способностью поглощать ультрафиолетовые лучи (защитная функция). Находящиеся в составе РВНСТ пигментоциты (меланофоры) сами не способны синтезировать меланин, они получают его от меланинн-родуцирующих клеток меланоцитов, которые находятся в составе эпителия (эпидермис и др.). Поэтому для структуры меланофоров в отличие от меланоцитов характерно слабое развитие органелл белкового синтеза. Адвентициальные клетки. Это малодифференцированные клетки с высоким ЯЦО, слабобазофильной, бедной органеллами цитоплазмой и большой способностью к митозу. Лежат возле гемокапилляров (поэтому их второе название — периваскулярные клетки, ПВК). Их считают стволовыми клетками для ФБ и липоцитов. Предшественники адвентициальных клеток мигрируют в РВНСТ из костного мозга, где имеется популяция самоподдерживающихся стволовых клеток для стромальных механоцитов.
ПЕРИЦИТЫ. Это клетки, окружающие сосуды микроциркуляторного русла, в первую очередь гемокапилляры. Некоторые авторы считают их предшественниками фибробластов. Подробнее о перицитах см. раздел частной гистологии "Сердечно-сосудистая система".
|
ЛЕЙКОЦИТЫ. Из крови в РВНСТ попадают все виды лейкоцитов: гранулоциты, лимфоциты, моноциты, последние превращаются в макрофаги (все лейкоциты — "пришлые" клетки РВНСТ).
МЕЖКЛЕТОЧНОЕ ВЕЩЕСТВО. Состоит из волокон и основного (аморфного) вещества (рис. 10.8). Волокна делятся на коллагеновые, элас-тические, ретикулярные Межклеточное вещество образуется клетками РВНСТ. Главны ми его продуцентами являются фибробласты, которые синтезируют компоненты как волокон, так и основного вещества. Тучные клетки также синтезируют некоторые компоненты основного вещества. Часть основного вещества образуется из плазмы крови.
КОЛЛАГЕНОВЫЕ ВОЛОКНА (рис. 10.8). Состоят из белка коллагена. В настоящее время описаны 19 типов коллагена, из которых наибольшее значение имеют пять:
— первый тип находится в соединительной ткани кожи, кости, стенке артерий;
— второй тип обнаружен в хрящевой ткани;
— третий тип встречается в дерме плода, в крупных сосудах, ретикулярных волокнах;
— четвертый тип входит в состав базальных мембран и капсулы хрусталика;
— пятый тип также участвует в образовании базальных мембран, а также стенки кровеносных сосудов, связок, дентина, основного вещества
роговицы. Коллагены I, II, III и V типов являются фибриллярными, т.к. способны формировать филаменты и фибриллы. Остальные коллагены этой способностью не обладают и являются аморфными. VI—XIX типы
коллагена мало изучены.
Молекулы коллагенов образованы тремя закрученными в виде спирали нитями ос-цепями. Специфичен аминокислотный состав цепей: в них преобладают аминокислоты глицин, пролин, лизин, гидроксинролин и i идроксилизин. Молекула коллагена синтезируется в ФБ. Помимо их, к коллагенсинтези-рующим клеткам относятся остеобласты, хондробласты, цементоблас-ты, дентинобласты, ретикулярные клетки, гладкие миоциты, клетки периневрия. Процессы биосинтеза коллагена во всех этих клетках похожи. Их можно разделить на два этана: внутриклеточный и внеклеточный.
Внутриклеточный этап. Вначале из аминокислот в гранулярной ЭПС образуются полипептидные ос-цени. Синтезированные пени накапливаются в цистернах гранулярной ЭПС и при этом подвергаются модификации: с участием витамина С происходит гликозилирование гидроксилизииа, а также образуются дисульфидные мостики. Недостаток витамина С приводит к образованию слабогидроксилированиых полипептидных цепей, не способных скручиваться в тройные спирали. Далее образуются молекулы проколлагена, состоящие из трех полипептидных цепей, сдвинутых одна по отношению к другой на 1/4 длины. В результате молекула коллагена и коллагеновые волокна имеют поперечную исчерченность. Далее молекулы проколлагена с помощью транспортных пузырьков поступают в комплекс Гольджи, где подвергаются терминальному гликозилироваиию. Молекулы проколлагена в комплексе Гольджи оформляются в секреторные гранулы и затем секретируются в межклеточное вещество.
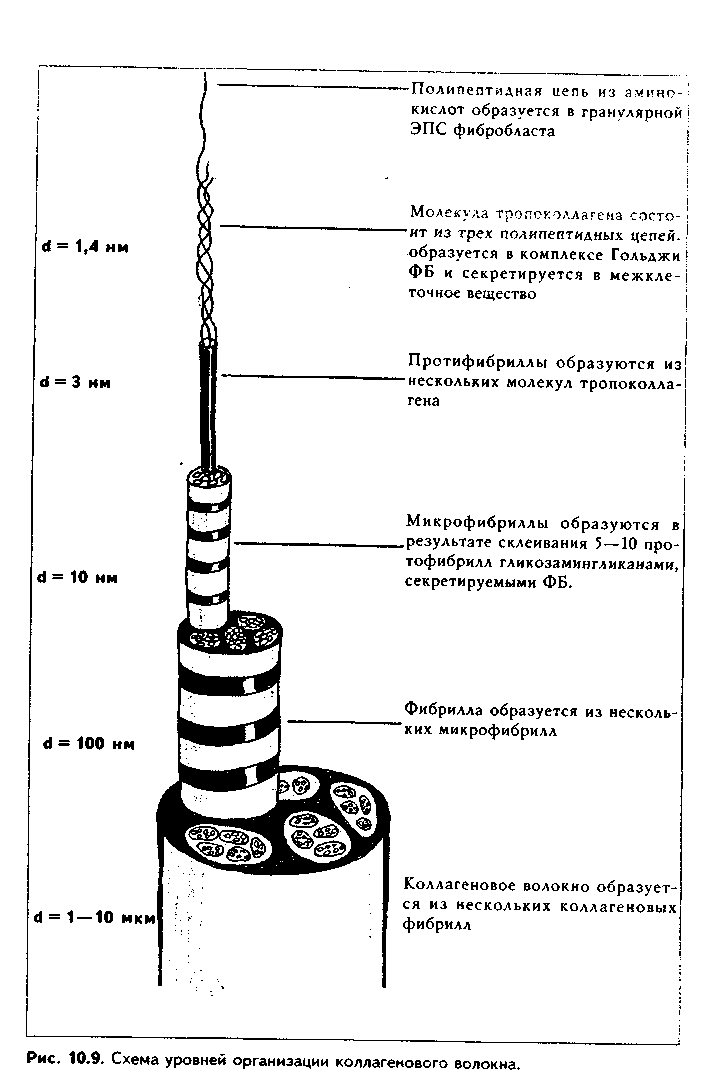
Внеклеточный этап. После секреции в межклеточное вещество с помощью пептидазы от молекул проколлагена отщепляются концевые участки с образованием молекул тропоколлагена. Этот процесс происходит только в фибриллярных коллагенах и приводит к появлению у них способности агрегировать в фибриллы. Далее происходит полимеризация молекул тропоколлагена: они последовательно соединяются конец в конец и сторона к стороне и образуют протофибриллы (1=3—5 им. Пять—шесть протофиб-рилл образуют микрофибриллы толщиной 10—20 им. Затем микрофибриллы склеиваются при помощи гликозаминогликаиов и гликопротеипов, секретируемых ФБ, образуя фибриллы толщиной 100 нм. Несколько фибрилл соединяются вместе и образуют видимые в световом микроскопе коллагеновые волокна толщиной 1—10 мкм (рис. 10.9). Таким образом, колла-геновое волокно имеет такие последовательные уровни организации: полипептидная цепь — молекула проколлагена — молекула тропоколлагена — протофибриллы — микрофибриллы — фибриллы — коллагеновое волокно.
В РВНСТ коллагеновые волокна образованы в основном коллагеном I типа. На гистологических препаратах они имеют вид оксифильных извитых тяжей, идущих в различных направлениях либо поодиночке, либо соединяясь в пучки различной толщины (см. рис.10.8). В поляризационном микроскопе коллагеновые волокна обладают двулучепреломлением, а в электронном микроскопе в них выявляются параллельно расположенные фибриллы с поперечной исчерченностыо (рис. 10.8).

|
Функция коллагеновых волокон: 1) опорная; 2) обеспечение прочности тканей; 3) информационно-регуляторная участие в морфойи
ноле, дифференцировке, регенерации клеток и тканей (в первую очередь, фибробластов), регуляции миграции, секреции и синтетической активности клеток, в адгезии клеток, а также тромбоцитов и образовании тромба; 4) участие в определении архитектоники соединительной ткани.
Эластичные волокна. Содержатся в РВНСТ в значительно меньшем количестве, чем коллагеновые. Состоят из аморфного эластина и образующего микрофибриллы фибриллина. Эластин и фибриллин синтезируются в гранулярной ЭПС, а затем модифицируются в комплексе Гольджи. Эластин, как и коллаген, содержит много глицина и пролина, а также две уникальные аминокислоты десмозин и изодесмозин. Молекулы эластина имеют вид глобул. После секреции в межклеточное вещество они соединяются в цепочки и образуют эластиновые протофибриллы толщиной 3 нм.
|
В последующем из протофибрилл в результате межмолекулярных связей образуется упругая резиноподобная сеть молекул, которая входит в так называемый светлый центральный аморфный компонент эластического волокна, расположенный в его центре. Снаружи от аморфного компонента, частично погружаясь в него, находится периферический фибриллярный компонент волокна, элементы которого образованы фибриллином. При образовании эластического волокна вначале из фибриллина образуются микрофибриллы (окситалановые волокна), которые служат основой для дальнейшего отложения эластина. Эластин постепенно накапливается в центральной части в форме аморфного компонента (элауниновые волокна), смещая фибриллярный компонент на периферию.
На светомикроскопическом уровне эластические волокна выявляются только при специальных окрасках (железный гематоксилин, орсеин и др., рис. 10.8). Имеют вид топких, прямых, часто ветвящихся и аиастомозиру-ющих между собой нитей, образующих трехмерную сеть. В силу своей молекулярной организации эластические волокна способны к возвращению в первоначальное состояние после растяжения — эластичности.
Помимо зрелых эластических волокон в РВНСТ всегда имеются незрелые эластические волокна: окситалановые и элауниновые. Окситалановые волокна образованы только микрофибриллами, формирующими периферический компонент волокна. В элаунииовых волокнах начинает формироваться центрально лежащий аморфный компонент, в котором еще содержатся микрофибриллы. В зрелых эластических волокнах фибриллярный компонент занимает только периферическое положение.
Кроме фибробластов эластические волокна синтезируют хондроблас-ты, хондроциты и гладкие миоциты.
Функции эластических волокон: 1) обеспечение обратимой деформации тканей — эластичности; 2) участие в создании архитектоники ткани.
Ретикулярные волокна. По своему химическому составу относятся к коллагеновым волокнам, т.к. состоят из белка коллагена (коллаген III тина). При обычной окраске гематоксилин-эозином не выявляются. Состоят из микрофибрилл, между которыми находятся цементирующие их гликонротеины и нротеогликаны. Благодаря их наличию ретикулярные волокна импрегнируют-ся солями серебра и дают положительную ШИК-реакцию (см. рис. 10.11). Ретикулярные волокна находятся в ретикулярной ткани кроветворных и имму-нокомпстентных органах, однако встречаются практически во всех видах соединительной ткани. К клеткам-продуцентам ретикулярных волокон кроме фибробластов относятся ретикулярные и жировые клетки, гладкие миоциты, кардиомиоциты, нейролеммоциты (см. Нервную ткань).
Основная функция ретикулярных волокон - опорная.
Аморфное вещество. Это второй компонент межклеточного вещества. При изучении в световом микроскопе прозрачно, может давать базофи-лию, в электронном микроскопе характеризуется низкой электронной плотностью.
Состоит из 90% воды, белков (сложные белки гликопротеины, протеог-ликаны, белки крови — альбумин, глобулин, фибриноген), жиров, углеводов (прежде всего, гликозаминогликаны), минеральных веществ. Молекулы гликозаминогликанов (гиалуроновая кислота и др.) имеют крупные размеры и формируют трехмерную сеть. В силу своей гидрофилыюсти они удерживают большое количество воды и формируют гель, через который диффундируют метаболиты. Гликопротеины и нротеогликаны обеспечивают взаимодействие межклеточного вещества с клетками, участвуют в транспорте воды и электролитов, аккумулируют ростовые факторы, выполняют ряд других функций. В зависимости от функционального состояния основное вещество может находиться в состоянии геля — коллоидное, и в состоянии золя — более жидкое состояние. Переход из состояния гель в состояние золь осуществляется ферментом гиалуронидазой, которая расщепляет гиалуроновую кислоту основного вещества. При этом снижается плотность основного вещества, увеличивается его проницаемость и проницаемость сосудов.
Функции аморфного вещества: регуляторная, обменно-трофическая, обеспечение микросреды для клеток.
|
|
|

Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
© cyberpedia.su 2017-2026 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

