Строение гена у эукариот.

Промоторы - связываются с РНК-полимеразой в процессе транскрипции.
-35 последовательность – именно с этой специф.-35 последовательностью оснований регуляторного участка гена связывается в начале сигма субьедицина РНК полимеразы.
Точка начала репликации или сайт «О» - последов. С которой начинается процесс репликации.
Энхансеры - увеличивают скорость транскрипции в 200 раз.
Сайленсеры - снижают скорость транскрипции.
Терминаторы – служат сигналом об окончании транскрипции (терминации).
ТАТА-бокс - модуль повт. в регуляторных элементах, служит в качестве сайта связ-я для РНК- полимеразы. Положение ТАТА-бокса строго определяет сайт инициации транскрипции, т.е. 5’ конец транскрипции.
ЦААТ-бокс - участвует в процессе инициации транскрипции.
ГЦ-бокс - влияет на эффективность транскрипции (скорость и процессивность).
Строение гена у прокариот.
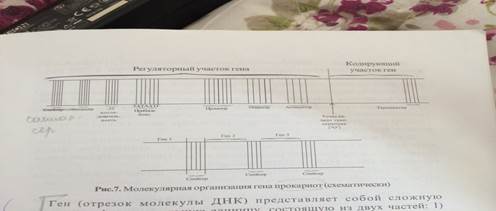
Промоторы - связываются с РНК-полимеразой в процессе транскрипции.
-35 последовательность – именно с этой специф.-35 последовательностью оснований регуляторного участка гена связывается в начале сигма субьедицина РНК полимеразы.
Прибнов-бокс - распознается и связывается РНК-полимеразы во время инициации транскрипции, яв-ся первой точкой, где комплемент цепи ДНК расходятся чтобы дать возможность начаться синтезу РНК на матрице ДНК.
Операторы - сайт связи с белками-репрессорами.
Точка начала репликации или сайт «О» - последов. С которой начинается процесс репликации.
Энхансеры - увеличивают скорость транскрипции в 200 раз.
Сайленсеры - снижают скорость транскрипции.
Аттенуаторы - запускают или предотвращают процесс транскрипции.
Терминаторы – служат сигналом об окончании транскрипции (терминации).
Репликация днк. Ферменты репликации
Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
1. инициация репликации
2. элонгация
3. терминация репликации.
ДНК – полимераза
ДНК-полимераза — фермент, участвующий в репликации ДНК. Ферменты этого класса катализируют полимеризацию дезоксирибонуклеотидов вдоль цепочки нуклеотидов ДНК, которую фермент «читает» и использует в качестве шаблона. Тип нового нуклеотида определяется по принципу комплементарности с шаблоном, с которого ведётся считывание. Собираемая молекула комплементарна шаблонной моноспирали и идентична второму компоненту двойной спирали.
Выделяют ДНК-зависимую ДНК-полимеразу, использующую в качестве матрицы одну из цепей ДНК, и РНК-зависимую ДНК-полимеразу, способную также к считыванию информации с РНК (обратная транскрипция).
ДНК-полимеразу считают холоферментом, поскольку для нормального функционирования она требует присутствия ионов магния в качестве кофактора. В отсутствии ионов магния о ней можно говорить как об апоферментe.
ДНК-полимераза начинает репликацию ДНК, связываясь с отрезком цепи нуклеотидов. Среднее количество нуклеотидов, присоединяемое ферментов ДНК-полимеразой за один акт связывания/диссоциации с матрицей, называют процессивностью.
ДНК – лигазы
Лигаза — фермент, катализирующий соединение двух молекул с образованием новой химической связи (лигирование). При этом обычно происходит отщепление (гидролиз) небольшой химической группы от одной из молекул.
Лигазы относятся к классу ферментов EC 6.
В молекулярной биологии лигазы разделяют на две большие группы — РНК-лигазы и ДНК-лигазы. ДНК-лигаза, осуществляющая репарацию ДНК
ДНК-лигазы — ферменты, катализирующие ковалентное сшивание цепей ДНК в дуплексе при репликации, репарации и рекомбинации. Они образуют фосфодиэфирные мостики между 5'-фосфорильной и 3'-гидроксильной группами соседних дезоксинуклеотидов в местах разрыва ДНК или между двумя молекулами ДНК. Для образования этих мостиков лигазы используют энергию гидролиза пирофосфорильной связи АТФ. Один из самых распространённых коммерчески доступных ферментов — ДНК-лигаза бактериофага Т4.
ДНК – геликазы
ДНК геликазы - ферменты раскручивающие двуцепочечную спираль ДНК с затратой энергии гидролиза трифосфатов NTP. Образуемая одноцепочечная ДНК участвует в различных процессах, таких как репликация, рекомбинация, и репарация. ДНК геликазы необходимы для репликации, репарации, рекомбинации и транскрипции. Геликазы присутствуют во всех организмах.
ДНК-топоизомеразы
ДНК-топоизомеразы—ферменты, изменяющие степень сверхспиральности и тип сверхспирали. Путём одноцепочечного разрыва они создают шарнир, вокруг которого нереплецированный дуплекс ДНК, находящейся перед вилкой, может свободно вращаться. Это снимает механическое напряжение, возникающее при раскручивании двух цепей в репликативной вилке, что является необходимым условием для её непрерывного движения. Кроме того, топоизомеразы (типа II) обеспечивают разделение или образование катенанов - сцепленных кольцевых ДНК (образуются в результате репликации кольцевой ДНК), а также устранение узлов и спутанных клубков из длинной линейной ДНК. Существует два типа топоизомераз. Топоизомеразы типа I уменьшают число сверхвитков в ДНК на единицу за один акт. Эти топоизомеразы надрезают одну из двух цепей, в результате чего фланкирующие дуплексные области могут повернутся вокруг интактной цепи, и затем воссоединяют концы разрезанной цепи. Эта реакция не требует энергии АТФ, т.к. энергия фосфодиэфирной связи сохраняется благодаря тому, что тирозиновый остаток в молекуле фермента выступает то в роли акцептора, то в роли донора фосфорильного конца разрезанной цепи.
Топоизомеразы типа II вносят временные разрывы в обе комплиментарные цепи, пропускают двухцепочечный сегмент той же самой или другой молекулы ДНК через разрыв, а затем соединяют разорванные концы. В результате за один акт снимаются два положительных или отрицательных сверхвитка. Топоизомеразы типа II тоже используют тирозиновые остатки для связывания 5¢-конца каждой разорванной цепи в то время. когда другой дуплекс проходит через место разрыва.
Праймаза
Праймаза—фермент, обладающий РНК-полимеразной активностью; служит для образования РНК-праймеров, необходимых для инициации синтеза ДНК в точке ori и дальнейшем для синтеза отстающей цепи.

7 Нарисовать и объяснить причины недорепликации, механизмы удлинения концов линейных молекул ДНК
Известно, что ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении от ее 3'-конца к 5'-концу. Соответственно дочерняя цепь синтезируется в направлении от 5' к 3'. В противоположном направлении синтез цепи ДНК фермент катализировать не может. Кроме того, ДНК-полимераза начинает синтез только со специального РНК-праймера - короткой РНК-затравки, комплементарной ДНК. После окончания синтеза ДНК РНК-праймеры удаляются, а пропуски в одной из дочерних цепей ДНК заполняются ДНК-полимеразой. Однако на 3'-конце ДНК такой пропуск заполнен быть не может, и поэтому 3'-концевые участки ДНК остаются однотяжевыми, а их 5'-концевые участки - недореплицированными. Отсюда ясно, что каждый цикл репликации хромосом будет приводить к их укорочению и прежде всего будет сокращаться длина теломерной ДНК.
Первым на проблему "концевой недорепликации ДНК" обратил внимание А.М. Оловников в 1971 году. Он высказал гипотезу о том, что потеря концевых последовательностей ДНК вследствие их недорепликации ведет к старению клетки. Иными словами, предполагалось, что процесс укорочения теломер и есть тот часовой механизм, который определяет репликативный потенциал "смертной" клетки, и когда длина теломер становится угрожающе короткой, этот механизм предотвращает дальнейшее деление клетки. А.М. Оловников предположил также, что в нестареющих клетках (а к ним кроме раковых относятся зародышевые, стволовые и другие генеративные клетки) должна существовать специализированная ферментативная система, которая контролирует и поддерживает длину теломерной ДНК.
Теломераза.
Теломераза — фермент, добавляющий особые повторяющиеся последовательности ДНК (TTAGGG у позвоночных) к 3'-концу цепи ДНК на участках теломер, которые располагаются на концах хромосом в эукариотических клетках. Теломеры содержат уплотненную ДНК и стабилизируют хромосомы.
Теломераза была впервые обнаружена в 1985 году у равноресничной инфузории Tetrahymena thermophila. Впоследствии она была обнаружена в дрожжах, растениях, животных, в.т.ч. яичниках человека и иммортализованных (бессмертных) линиях раковых клеток Hela.
Теломераза является ДНК-полимеразой, достраивающей 3'-концы линейных молекул ДНК хромосом короткими(6-8 нуклеотидов), повторяющимися последовательностями(у позвоночных она представлена нуклеотидами ТТАГГГ). Теломераза называется также ДНК-нуклеотидилэкзотрансферазой или теломерной терминальной трансферазой. Она состоит из белков и РНК, выполняющей роль матрицы для удлинения молекулы ДНК повторами нуклеотидов. Длина теломеразной РНК составляет от 150(простейшие) до 1400 нуклеотидов у дрожжей. У человека ее протяженность составляет 450 нуклеотидов.
Принцип действия теломеразы основан на удлинении не вновь синтезированной дочерней цепи, а материнской цепи ДНК. Процесс удлинения(достраивания) материнской(матричной) цепи ДНК теломеразой происходит до того, как эндонуклеазы отщепляет концевые нуклеотиды дочерней цепи.
По составу теломерных повторов в теломерах ДНК различают Г-цепь (гуаниновая цепь) и Ц-цепь (цитозиновая цепь). Обе цепи входят в состав двуцепочной молекулы ДНК. Теломераза удлиняет Г-цепь каждой теломеры.
РНК, содержащаяся в составе теломеразы, имеет матричный участок с помощью которого теломераза распознает теломерный повтор. После распознавания теломеры и связывания теломеразы с молекулрй ДНК последовательность оснований в составе молекулы полимеразы: 5'-ЦААЦЦЦАА-3' синтезирует последовательность, комплиментарную с последовательностью в теломерном повторе: 5'-ТТГГГГ-3'. Нуклеотиды ААЦ в РНК теломеразы остаются неспаренными, к ним присоединяются нуклеотиды ТТГ молекулы ДНК. Данный процесс осуществляется путем перемещения теломеразы на самый конец теломерной последовательности ДНК. В результате этого конец материнской ДНК удлиняется на повтор нуклеотидов-ТТГГГГТТГ.
Процесс образования нового теломерного повтора носит название элонгации.
Образование следующего теломерного повтора происходит путём перемещения теломеразы(вместе с РНК) на один повтор к 3'-концу удлиняемой цепи ДНК. Процесс перемещения теломеразы носит название транслокации. Таким образом процесс наращивания теломерного участка материнской хромосомы включает в себя этапы элонгации и транслокации теломеразы.
В конечном итоге теломераза может удлинить цепь ДНК на несколько десятков или сотен теломерных повторов(количество повторов зависит по-видимому от видовой принадлежности организма и количества делений,пройденных той или иной клеткой).
На удлиненной части ДНК достраивается дочерняя цепь ДНК. Мутации в теломерной РНК приводят не к синтезу ошибочных нуклеотидов, а к утрате активности всей теломеразы.
Следует особо подчеркнуть, что теломераза функционирует и восстанавливает дочернюю молекулу ДНК только в генеративных, раковых и линиях иммортализованных(бессмертных) клеточных культурах.
В соматических клетках теломеразная активность отсутствует и, следовательно, теломеры соматических клеток постепенно, после каждой репликации, укорачиваются.
В этой связи было высказано предположение, что постепенное укорочение ДНК хромосом в процессе репликации, может лежать в основе ограниченного числа делений клеток, культивируемых in vitro. Данное ограничение носит название «лимита Хейфлика». Хейфлик показал в начале 60-х годов XX века зависимость числа клеточных делений от возраста человека. У новорожденных соматические клетки могут делиться 80-90 раз, а у 70-летних людей- только 20-30 раз.
Укорочение теломерных концов хромосом можно считать молекулярным индикатором количества делений, но не старения клетки.
Прекращение работы теломеразы, наблюдаемое в большинстве дифференцированных соматических клеток, является свидетельством их зрелости и одновременно специфическим сигналом для запуска процессов, приводящих клетки к естественной гибели(апоптозу). 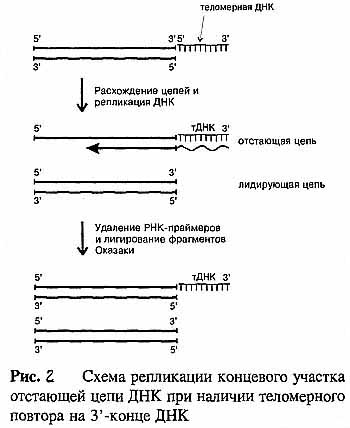
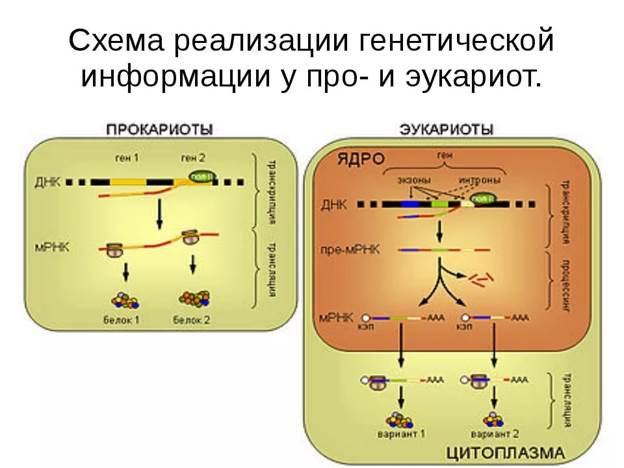
1. 8 9 Нарисовать, дать определение процессу транскрипции, охарактеризовать стадии транскрипции, строение и функции ферментов, обеспечивающих процесс транскрипции прокариот
ДНК-зависимая РНК-полимераза — это фермент, катализирующий синтез РНК. Он состоит из несколько субъединиц: двух α, одной β, одной β' и одной σ. Их комплекс называется холоферментом (α2ββ'σ) и имеет мол. массу (Мг) около 500 000. Фермент, лишенный σ-субъединицы, называется кор-ферментом. Для инициации транскрипции необходимы холо-фермент, нуклеозидтрифосфат (всегда АТР или GTP) и наличие специального участка в ДНК, называемого промотором. Когда полимераза связывается с промотором, происходит локальное расплетание двойной спирали ДНК и образуется открытый промоторный комплекс. Промотор - это участок молекулы ДНК, имеющий размер около 40 пар оснований и расположенный непосредственно перед участком инициации транскрипции. Синтез РНК всегда начинается с оснований А или G в «+»-цепи ДНК. Участок связывания холофермента расположен «левее» сайта инициации (в направлении 3' - 5' в «+»-цепи) на расстоянии примерно 10 оснований. Если сравнить последовательности оснований «+»-цепи ДНК у разных промоторов, то мы обнаружим, что они весьма близки, хотя и не идентичны. Эта так называемая последовательность Прибнова имеет вид TATPuATPu, где Рu означает пурин (А или G). Таким образом, холофермент связывается со специфической последовательностью или группой последовательностей. Обычно на расстоянии около 40 оснований «левее» участка инициации находится второе место связывания РНК-полимеразы, где, как полагают, происходит связывание а-субъединицы с ДНК.
Элонгация цепи РНК — это та стадия транскрипции, которая наступает после присоединения примерно восьми рибонуклеотидов. В этот момент РНК- полимераза претерпевает структурное изменение, при котором от комплекса отделяется σ-субъединица и остается кор-фермент (α2ββ'), катализирующий дальнейшее удлинение цепи РНК. При этом к цепи присоединяются те рибонуклеозидтрифосфаты, которые обеспечивают правильное спаривание с «—»-цепью ДНК. Движущийся вдоль ДНК кор-фермент действует подобно застежке-молнии, «раскрывая» двойную спираль, которая замыкается позади фермента по мере того, как соответствующие основания РНК спариваются с основаниями ДНК в «—»-цепи. «Раскрытая» ферментом область простирается только на несколько пар оснований.
Терминация (прекращение роста) цепи РНК происходит на специфических участках ДНК, называемых терминаторами. Начало этих участков обычно обогащено GC-парами, а остальная последовательность — АТ-парами. GC-богатый участок часто представляет собой палиндром. Это означает, что при движении вдоль «+»-цепи в одном направлении, а вдоль «—»-цепи - в противоположном читается одна и та же последовательность оснований. В остановке синтеза РНК именно на терминаторе важную роль играет р-белок.
^ Посттранскрипционный процессинг — это процесс созревания, при котором первичный РНК-транскрипт модифицируется и превращается в зрелую РНК. Характер и степень модификации РНК зависят от типа РНК.
Молекулы мРНК у прокариот не подвергаются процессингу. У некоторых бактерий транскрипция и трансляция сопряжены, т. е. происходят одновременно. 5'-конец мРНК может транслироваться на рибосоме и затем подвергаться деградации еще до завершения синтеза ее 3'-конца. Молекулы тРНК вначале синтезируются в виде про-тРНК, которая примерно на 20% длиннее, чем соответствующая тРНК. Лишние последовательности, расположенные у 5'- и 3'- концов, удаляются с помощью таких ферментов, как рибонуклеазы Q и Р. Иногда молекула про-тРНК состоит из двух или более молекул тРНК, соединенных между собой. Их разделение также осуществляется с помощью рибонуклеаз. Если 3'-конец тРНК не несет концевой последовательности ССА, то эти основания присоединяются при постсинтетической модификации. Все тРНК содержат минорные основания, которые являются химически модифицированными формами четырех главных оснований (А, С, G и U).
Эта модификация происходит после завершения транскрипции. Гены рРНК прокариот расположены в транскрипционных блоках. Три генарРНК Е. coli (16S, 23S и 5S) располагаются вместе с генами нескольких тРНК в одном таком блоке и транскрибируются в виде одной молекулы РНК. Эти молекулы рРНК и тРНК отделены друг от друга спейсерной РНК. Расщепление первичного транскрипта на отдельные составляющие катализирует рибонуклеаза Q; поскольку этот фермент специфичен к двухцепочечной РНК, предполагают, что в области спейсеров образуются двухцепочечные шпильки, которые фермент узнает и вырезает.
^ 3. Транскрипция у эукариот
У эукариот для транскрипции используются три ДНК - зависимых РНК-полимеразы. Полимераза I локализована в ядрышке, где она катализирует синтез рРНК в виде большого первичного транскрипта, содержащего молекулы рРНК 18S, 5,8S и 28S. Полимераза II находится в нуклеоплазме и, вероятно, участвует в синтезе первичного транскрипта мРНК. Полимераза III также локализована в нуклеоплазме и участвует в синтезе тРНК и 5S-pPHK.
Синтез РНК включает стадии инициации, элонгации и терминации, но в этих процессах часто принимают участие другие ферменты и последовательности оснований, чем у прокариот. Например, промоторные последовательности у эукариот отличаются от таковых у прокариот. Однако первыми основаниями, включаемыми в РНК при инициации, являются, как и у прокариот, А или G.
Молекулы мРНК обычно образуются из больших по размеру молекул-предшественников, называемых гетерогенной ядерной РНК (гяРНК). Для образования зрелой мРНК эти молекулы подвергаются модификации по 5'- и З'-концами и сплайсингу. После такой модификации транскрипты переносятся из ядра в цитоплазму.
Сплайсинг гяРНК — это удаление последовательностей РНК, соответствующих интронам ДНК, и соединение участков, которые транскрибированы с кодирующих последовательностей (экзонов). Место сплайсинга должно быть определено с высокой точностью, поскольку ошибка даже в одно основание приведет к синтезу белка с неправильной аминокислотной последовательностью. Такая специфичность сплайсинга обеспечивается строго определенной последовательностью оснований в интроне, отвечающей обычно основаниям GU или GA в начале соответствующей РНК и основаниям AG - в конце.
Модификация 5'-конца мРНК приводит к образованию особой последовательности, называемой кэп-структурой. При модификации 3'-конца к нему присоединяется последовательность poly(A) длиной 150-200 нуклеотидов.
Процессинг тРНК у эукариот протекает по такому же механизму, как и у прокариот. Функционально активные молекулы образуются из более длинного предшественника, который подвергается расщеплению и модификации с включением минорных оснований.
Процессинг рРНК также аналогичен соответствующему процессу у прокариот. Первичный транскрипт содержит участки, отвечающие 18S-, 5,8S- и 28S-рРНК, разделенные спейсерами. Как и у прокариот, эти три рРНК образуются при расщеплении спейсерных последовательностей.
2. 10 Нарисовать и объяснить процессы созревания ядерной и-РНК эукариот
11 Рибонуклеиновая кислота (РНК) – это однонитевой биополимер, в качестве мономеров которого выступают нуклеотиды.
Матрицей для синтеза новых молекул РНК являются молекулы дезоксирибонуклеиновой кислоты (транскрипция РНК). Хотя в ряде случаев возможен и обратный процесс (образование новых ДНК на матрице РНК в ходе репликации некоторых вирусов). Также основой для биосинтеза РНК могут быть другие молекулы рибонуклеиновой кислоты (репликация РНК). В транскрипции РНК, происходящей в ядре клетки, участвует целый ряд ферментов, наиболее значимым из которых является РНК-полимераза.
Строение РНК
Молекула имеет однонитевое строение. Полимер. В результате взаимодействия нуклеотидов друг с другом молекула РНК приобретает вторичную структуру, различной формы (спираль, глобула и т.д.). Мономером РНК является нуклеотид (молекула, в состав которой входит азотистое основание, остаток фосфорной кислоты и сахар (пептоза)). РНК напоминает по своему строению одну цепь ДНК.
Нуклеотиды, входящие в состав РНК: гуанин, аденин, цитозин, урацил. Аденин и гуанин относятся к пуриновым основаниям, цитозин и урацил к пиримидиновым. В отличие от молекулы ДНК, в качестве углеводного компонента рибонуклеиновой кислоты выступает не дезоксирибоза, а рибоза. Вторым существенным отличием в химическом строении РНК от ДНК является отсутствие в молекуле рибонуклеиновой кислоты такого нуклеотида как тимин. В РНК он заменён на урацил.
Сцепленное наследование
Если гены А и В расположены в
· одной хромосоме;
· одной паре хромосом;
· одной паре гомологичных хромосом;
то они не смогут разойтись независимо, происходит сцепленное наследование согласно закону сцепления Моргана: "Гены, расположенные в одной хромосоме, наследуются совместно".
Все гены, находящиеся в одной хромосоме, образуют группу сцепления. Количество групп сцепления равняется количеству хромосом в гаплоидном наборе (количеству пар гомологичных хромосом)
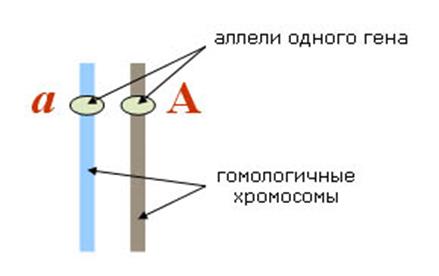
7) АЛЛЕЛЬНЫЕ ГЕНЫ — это гены, занимающие одинаковые локусы в гомологичных хромосомах и отвечающие за развитие альтернативных признаков. Аллельные гены, или аллели, представляют собой разные формы одного и того же гена.
Аутосомный синром
Синдром Дауна. Первое клиническое описание синдрома Дауна относится к 1866г. Частота 1: 550 - 650 новорожденных. Среди умственно отсталых детей выявляется 10-12% больных с синдромом Дауна.
При цитогенетическом анализе выявляют 3 формы:
- простая трисомия по хромосоме 21 (в 95% случаев)
- транслокация хромосомы 21 на другие хромосомы (чаще на 14, 15, реже на 21, 22 и Y-хромосому) (4% случаев)
- мозаичный вариант синдрома (1 %).
Больные с-м Дауна имеют характерный фенотип: брахицефалическая форма черепа с укорочением передне-заднего размера и утолщением затылка; раскрытые роднички, избыток кожи на затылке, плоский профиль лица, характерный разрез глаз, помутнение хрусталика, нистагм, короткий нос с широким переносьем, полуоткрытый рот с толстыми губами и большим языком, Руки короткие и широкие, клинодактилия мизинцев, часто наблюдаются врожденные пороки сердца, деформации скелета, задержка психомоторного развития, с возрастом нарастает интеллектуальный дефицит.
При транслокации симптоматика более выражена, чем при обычной трисомии. В кариотипе обнаруживают 46 хромосом, одну обычную хромосому 21 и одну маркерную хромосому, включающую хромосому 21, транслоцированную на другую хромосому.
Анализ кариотипов родителей детей с транслокационной формой синдрома нередко выявляет у фенотипически нормальной матери пробанда 45 неизмененных хромосом и наличие маркерной хромосомы - точно такой же, как у её больного ребенка.
При мозаичной форме синдрома выраженность фенотипа зависит от доли трисомных клеток в аномальном кариотипе пробанда: чем их больше, тем более выражен фенотип синдрома.
Синдром Патау. Эта трисомия по хромосоме 13 встречается с частотой 1: 6000 новорожденных. Цитогенетически различают 3 формы: трисомная (75%), транслокационная (20%) и мозаичная (5%). Фенотип включает в себя прежде всего, триаду признаков: микрофтальм, расщелины губы и неба, полидактилия. Черепно-лицевой дисморфизм характеризуется также микроцефалией, черепом неправильной формы, скошенным узким лбом, раскрытыми родничками и швами черепа, дефектами скальпа, узкими горизонтальными щелями. Нос плоский,широкий, переносье запавшее. Ушные раковины расположены низко.
При с-ме Патау центральная нервная система поражена во всех случаях. Характерно множественное поражение внутренних органов: пороки сердца, поликистоз почек, аномалии желудочно-кишечного тракта. Продолжительность жизни больных резко снижена. Обычно дети погибают в первые дни или недели жизни.
Синдром «кошачьего крика». Синдром делеции часто короткого плеча хромосомы 5. Частота синдрома - 1: 650 000; среди детей с задержкой умственного развития - 1: 350. Описаны мозаичные формы.
Дети с этим синдромом, как правило, рождаются после нормально протекавшей беременности. В неонатальном периоде состояние резко ухудшается: приступы посинения, удлиненный выдох, снижение двигательной активности, угнетение сосательного рефлекса, рвота.
Фенотип характеризуется монотонным или резким криком, похожим на кошачье мяуканье. С возрастом крик исчезает. Выявляется грубое физическое и интеллектуальное недоразвитие. Выражен черепно-лицевой дисморфизм (особенно в первые 2 года жизни): микроцефалия, вытянутая форма черепа, лунообразное лицо, узкие глазные щели, вдавленное переносье. Зубы расположены неправильно, передние резцы выступают вперед, рот широкий, толстая нижняя губа. Часто наблюдаются врожденные пороки сердца, почек, косолапость, сращение 2 и 3-го пальцев ног и некоторые другие признаки
45. Характеристика гоносомных синдромов Гоносомные синдромы подразделяются на синдромы с мужским и женским фенотипами, а также на клинические формы гермафродитизма.
Полисомия по У - хромосоме. Это хромосомный синдром с мужским фенотипом, общее название - с-м Клайнфельтера. Классическая форма - 47, ХХУ, составляющая 80% всех случаев синдрома, частота в популяции 1: 500, на оставшиеся 20% приходятся 48 ХХХУ и варианты мозаичной формы синдрома. Общим для всех вариантов и форм является триада признаков: гинекомастия, атрофия и гипоплазия тестикул и бесплодие. Вследствие гормонального дисбаланса такие больные имеют высокий рост, узкие плечи, широкий таз, скудное оволосение. Уровень гонадотропинов в моче соответствует норме или повышен.
Синдром полной моносомии по Х-хромосоме. Это синдром с женским фенотипом. Единственным примером этого синдрома является с-м Шерешевского - Тернера. Частота синдрома 1: 1430 у новорожденных девочек. Классический вариант - 45, ХО (55% всех случаев). Возможны варианты:
- мозаики 45, ХО/46, ХХ; 45, ХО/47, ХХХ
- делеции: 45, ХО, del (Х)
- кольцевая хромосома: 45, ХО, r (Х)
- другие варианты.
Фенотипически это лица женского пола с задержкой роста и полового развития. У них нормальные наружные и недоразвитые внутренние половые органы. Характерный признак - короткая шея с низкой линией роста волос на затылке и крыловидными складками кожи на шее.
30) Нарисрвать и объяснить причины и механизмы возникновения моносомий и трисомий у человека Синдром полной моносомии по Х-хромосоме. Это синдром с женским фенотипом. Единственным примером этого синдрома является с-м Шерешевского - Тернера. Частота синдрома 1: 1430 у новорожденных девочек. Классический вариант - 45, ХО (55% всех случаев). Возможны варианты:
- мозаики 45, ХО/46, ХХ; 45, ХО/47, ХХХ
- делеции: 45, ХО, del (Х)
- кольцевая хромосома: 45, ХО, r (Х)
- другие варианты.
Фенотипически это лица женского пола с задержкой роста и полового развития. У них нормальные наружные и недоразвитые внутренние половые органы. Характерный признак - короткая шея с низкой линией роста волос на затылке и крыловидными складками кожи на шее.
Трисомия
Трисомия — это наличие трёх гомологичных хромосом вместо пары (в норме). Причиной подавляющего большинства трисомий у человека являются ошибки расхождения хромосом при оогенезе, причём наибольший вклад дают ошибки в мейозе I по сравнению со вторым мейотическим делением. Вероятность трисомий у потомства повышается с возрастом матери. Единственной жизнеспособной трисомией по аутосоме у человека является трисомия по хромосоме 21, вызывающая синдром Дауна. Трисомики по хромосомам 13 (синдром Патау) и 18 (синдром Эдвардса) могут дожить до рождения
35) Основным свойством раковой клетки является ее безудержное и неконтролируемое деление, обусловленное отсутствием тормозящих пролиферацию клеток механизмов и факторов. Злокачественные опухоли являются по происхождению моноклональными, т е все клетки опухоли происходят от одной-единственной клетки – предшественницы и являются ее потомками. Автономность. Все злокачественные опухоли растут "сами по себе", они не подчиняются, не реагируют на различные (нервные, гуморальные) сигналы организма-хозяина. Клетки такой опухоли практически полностью утрачивают зависимость от внешних и внутренних факторов, они представлены сами себе. Склонность к проникновению («инвазии», «инфильтрации», «пенетрации») в окружающие ткани, с формированием местных метастазов.
Склонность к метастазированию в другие, часто весьма отдалённые от исходной опухоли ткани и органы посредством перемещения по лимфо- и кровеносным сосудам, а также имплантационно. Причём определённые типы опухолей проявляют определённое родство («тропность») к определённым тканям и органам — метастазируют в определённые места (но могут метастазировать и в другие).
34) Существует определенный класс генов, вызывающих злокачественную трансформацию клеток. Эти гены называются онкогенами. Протоонкогенами называются нормальные аналоги или предшественники окогенов.Превращение протоокогенов в онкогены осущ путем:
1) присоедеинения к протоокогену нового транскрипционного промотора
2) амплификация(увеличения числа копий) протоонкогена до онкогенной активности
3) включения в молекулу ДНК клетки хозяина некоторой последовательности нуклеотидов, усиливающей действие промотора
4) присоединение протоонкогена к локусу иммуноглобулина в результате транслокации
5) мутации протоонкогена
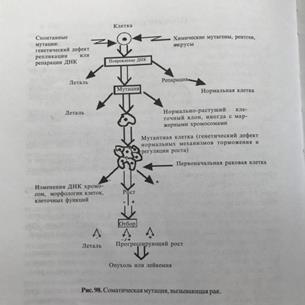
33) Живые организмы и их клетки не вечны: рано или поздно их существование заканчивается гибелью. Это естественный, генетический запрограммированный процесс, получивший название апоптоза. В общем виде процесс апоптоза можно представить следующим образом
Нарушение структуры и функционирования клеток – апоптоз - гибель клеток – элиминация.
В процессе апоптоза принимают участия специфические ферменты каспазы (бывают сериновые и цистеиновые) В нормальных клетках они представлены неактивными предшественниками прокаспазами. Они активируется в случае нарушение структуры клеток.
Апоптоз по команде: вызывается внешней сигнализацией, которая передается через мембранные рецепторы. Вызывается если отсутствует один из факторов:1) ростовой фактор 2)прикрепление 3) контактное торможение. Первым в ответ на сигнал активируется К8. Сигнал о нарушениях структур клетки может поступать также из митохондрий. Сигнальным фактором в этом случае является протеаза AIF. Он способствует превращению неактивной формы прокаспазы 9 в активную каспазу 9. В процессе активизации прокаспазы 9 принимает участие Цитохром С, стимулирующий взаимодействие прокаспазы 9 с белком Apaf 1 т другими прокаспазами 9. Ключевым ферментом апоптоза является каспаза 3, активируемая каспазами 8 и 9. Каспаза 3 активирует другие каспазы 6 и 7, и некоторые другие некаспазные белки.
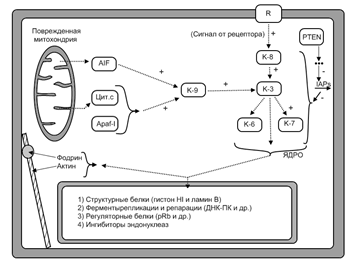
Апоптоз изнутри Может происходит из за нездорового состояния клетки, чрезмерные повреждений хромосом, повреждение внутриклеточной мембраны. Белок р53 является одним из ключевых белков апоптоза. В нормальной клетке количество и активность белка р53 поддерживается на минимальном уровне. Концентрация и активность белка р53 контролируется белком Mdm2, который при взаимодействии с белком с белком р53 снижает его активность и ускоряет его распад. Избыток белка р53 приводит к активации белка Mdm2 с последующим снижением активности и ускорением распада белка р53. Регуляторный фактор белок Arf или 19Arf задерживает распад белка р53. Еще один регуляторный белок 14-3-3в служит активатором белка р 53. Высокая активность белка р 53 в то время как белок Mdm2 регулирует как концетрацию так и активность белка р53.

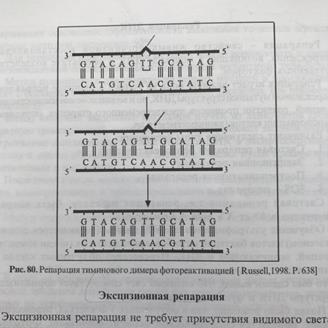
32). Пострепликативная репарация Наименее изученный тип репарации. Пострепликативная репарация осуществляется путем рекомбинации (обмена фрагментами) между двумя вновь образованными двойными спиралями ДНК. Примером такой пострепликативной репарации может служить восстановление нормальной структуры ДНК при возникновении тиминовыхдимеров (Т-Т), когда они не устраняются самопроизвольно под действием видимого света (световая репарация) или в ходе дорепликативнойэксцизионной репарации.
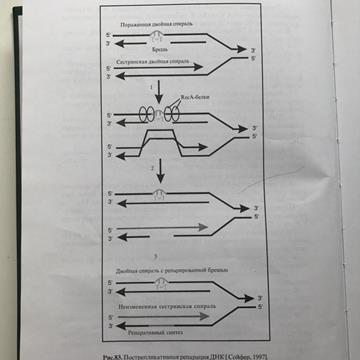
Важным механизмом удаления повреждений в ДНК является эксцизионная репарация (1).Специфическая нуклеаза удаляет небольшой сегмент ДНК, включающий поврежденный участок. Удаленный участок восстанавливается ДНК-полимеразой, использующей в качестве матрицы комплементарную цепь. Наконец, оставшийся одноцепочечный разрыв закрывается ДНК-лигазой.Тиминовыедимеры могут быть удалены фотореактивацией (2). Специфическая фотолиаза связывается с дефектным участком ДНК и после облучения расщепляет димер с образованием отдельных нуклеиновых оснований. Третий механизм — это репарация в результате рекомбинации (3, показано в упрощенном виде). В этом процессе участок, содержащий повреждение, пропускается во время репликации. Образующаяся брешь закрывается путем сдвига соответствующего сегмента из правильно реплицированной второй цепи. Новая брешь ликвидируется с участием полимераз и ДНК-лигаз. В завершение первоначальный дефект (1) устраняется путем вырезания.
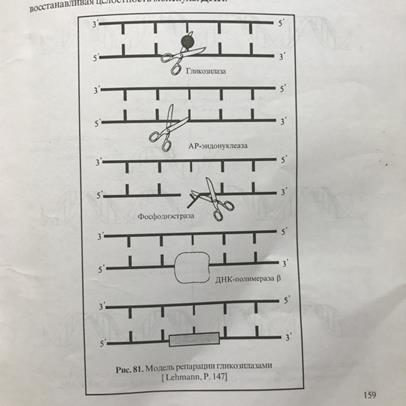
31) 1.Световая репарация. Механизм это репарации заключается в том, что под действием ультрафиолетовых лучей, в молекуле ДНК могут возникать сцепления между соседними пиримидиновыми основаниями с образованием димеров – Т=Т, Т=Ц, Ц=Ц, Ц=У, Т=У, У=У. квант видимого света активизирует специальный фермент, которые соединяются с поврежденной ДНК, разъединяют димеры и восстанавливает целостность нити ДНК.
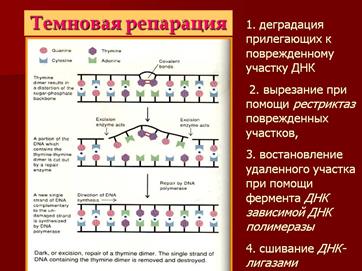
2.Темновая репарация. Клетки могут ликвидировать повреждения ДНК и без участия света. Стадии: а)узнавание повреждения ДНК, при помощи фермента нуклеазы б)вырезания, с помощью экзонуклеазы в) матричный синтез новой цепи с помощью полимераз г) соединение новообразованного участка.
19. Клинико-генеалогический метод включает клиническое обследование членов семьи пациента, обратившегося за консультацией, составление ее родословной и проведение генеалогического анализа. Генеалогический анализ является самым распространенным, наиболее простым и одновременно высоко информативным методом, доступным каждому, кто интересуется своей родословной и историей своей семьи. Он не требует ника






