

Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Топ:
Эволюция кровеносной системы позвоночных животных: Биологическая эволюция – необратимый процесс исторического развития живой природы...
Устройство и оснащение процедурного кабинета: Решающая роль в обеспечении правильного лечения пациентов отводится процедурной медсестре...
Характеристика АТП и сварочно-жестяницкого участка: Транспорт в настоящее время является одной из важнейших отраслей народного хозяйства...
Интересное:
Подходы к решению темы фильма: Существует три основных типа исторического фильма, имеющих между собой много общего...
Национальное богатство страны и его составляющие: для оценки элементов национального богатства используются...
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
В формировании признаков пола выделяют четыре уровня:
- хромосомное определение пола;
- определение пола на уровне гонад;
- фенотипическое определение пола (половых признаков);
- психологическое определение пола.
Хромосомное определение пола у животных и человека происходит в момент оплодотворения. Для человека это формирование кариотипа 46 XX или 46 ХУ, что определяется гаметой гетерогаметного пола. У человека женский пол гомогаметный, а мужской пол гетерогаметный. У птиц и бабочек, наоборот, самцы гомогаметные, а самки - гетерогаметные. У прямокрылых насекомых самки гомогаметны, с кариотип XX, а самцы гетерогаметны - ХО, у последних отсутствует у-хромосома.
Определение пола на уровне гонад у человека начинается с того, что на 3 - й неделе эмбрионального развития в энтодерме желточного мешка появляются первичные зародышевые клетки, которые под действием хемотаксических сигналов мигрируют в область закладки гонад (половых желез). Дальнейшее развитие признаков пола определяется наличием или отсутствием в кариотипе у-хромосомы.
Семенники развиваются, если имеется Y-хромосома. Под контролем у-хромосомы в первичных зародышевых клетках начинает синтезироваться Н-Y-антиген, который кодируется структурным аутосомным геном, контролируемым Y-хромосомой. Для превращения зачатка гонады в семенник достаточно уже малой концентрации Н-Y-антигена. На развитие семенников также оказывает влияние, по меньшей мере, ещё 19 генов: аутосомных и сцепленных с Х-хромосомой. А под действием хориогонического гонадотропина, секретируемого плацентой матери, в семенниках начинают вырабатываться мужские половые гормоны (андрогены) - это тестостерон и 5-дигидро-тестостерон.
Фенотипическое определение пола в виде развития внутренних и наружных половых органов и развития всего фенотипа по мужскому типу происходит следующим образом. Сцепленный с X-хромосомой ген (Tfm+) кодирует белок-рецептор, который, связываясь с тестостероном, доставляет его в ядра клеток, где тестостерон активизирует гены, обеспечивающие дифференцировку развивающегося организма по мужскому типу, в том числе и развитие семявыносящих путей. У зародыша человека из протока первичной почки формируются два протока: мюллеров и вольфов. У мужчин редуцируются мюллеровы протоки, а вольфовы преобразуются в семенные протоки и семенные пузырьки. При мутации гена Tfm+ и дефекте, рецепторов тестостерона может развиться синдром тестикулярной феминизации. В таких случаях у лиц с мужским кариотипом наружные половые органы развиваются по женскому типу. При этом влагалище бывает укорочено и заканчивается слепым мешком, а матка и маточных трубы отсутствуют. По пропорциям тела такие женщины приближаются к типу манекенщиц. Отмечается аменорея (отсутствие менструаций). В то же время молочные железы развиты нормально. Психологическое развитие у них осуществляется по женскому типу, хотя имеет место мужской кариотип и вместо яичников у них присутствуют семенники, которые располагаются либо в больших половых губах, либо в паховом канале, либо в брюшной полости. Сперматогенез отсутствует.
Рецепторы к гормонам имеют не только клетки-мишени тех или иных половых органов, но и нейроны головного мозга. Влияние гормонов на головной мозг начинается уже в эмбриональном периоде, что сказывается в дальнейшем и на особенностях сексуального поведения.
Если в кариотипе зиготы отсутствует У - хромосома, формируется женский фенотип без участия специальных регуляторных факторов. При этом из двух протоков, формирующихся из протока первичной почки, вольфов проток редуцируется, а мюллеровы преобразуются в матку и маточные трубы.
Сцепленное наследование, кроссинговер, определение расстояния между генами на эксперименте с дрозофилами. Группы сцепления, карты хромосом.
СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ
Г. Мендель изучил наследование только семи пар признаков у душистого горошка. Его законы подтвердились на самых разных видах организмов, т. е. было признано, что эти законы носят всеобщий характер. Однако позже, в 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Постепенно таких исключений из третьего закона Менделя накапливалось все больше. Стало ясно, что принцип независимого распределения в потомстве и свободного комбинирования распространяется не на все гены.
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы изучено более 500 генов, у мухи дрозофилы – более 1 тыс., а у человека – около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно.
Это дало основание предположить, что в каждой хромосоме локализовано множество генов.
Экспериментальные исследования явления сцепленного наследования провел выдающийся американский генетик Т. X. Морган со своими сотрудниками. Сцепленным наследованием называется такой вариант наследования признаков, при котором гены, отвечающие за развитие данных признаков, располагаются в одной хромосоме, составляют группу сцепления (рис.7) и наследуются совместно (сцепленно).
Если Г. Мендель проводил свои опыты на горохе, то для Т. Моргана основным объектом стала плодовая мушка дрозофила..
Дрозофила каждые две недели при температуре 25°С дает многочисленное потомство. Самец и самка внешне хорошо различимы – у самца брюшко меньше и темнее (рис. 8). Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
Томас Морган, как в свое время Грегор Мендель в опытах с горохом, изучал наследование 2 пар признаков: окраска тела и длина крыльев у мушки дрозофилы. Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении (F1) Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, – над геном недоразвитых).
Следовательно, в F1 Томас Морган получил единообразие гибридов первого поколения, как и Грегор Мендель, но в F2 Томас Морган никогда не получал соотношения 9:3:3:1 как в третьем законе Менделя. Тогда для выяснения генотипов особей первого поколения (F1) он провел анализирующее скрещивание.
Анализирующее скрещивание проводится с целью установления генотипа исследуемой особи. Для этого исследуемую особь (?) скрещивают с рецессивной гомозиготой (аа).
Если в F1 наблюдается расщепление 1:1, то исследуемая особь является по генотипу гетерозиготой – Аа.
| P: ♀ (?)Х ♂ аа G:?a F1: Aa; aa – 1:1 |
| P: ♀ AaХ ♂ аа G: A, a a F1: Aa; aa – 1:1 |
| выяснение генотипа |
Если в F1 наблюдается единообразие, то исследуемая особь является по генотипу гомозиготой – АА или аа.
| P: ♀ (?)Х ♂ аа G:?a F1: Aа – 100 % |
| P: ♀ AAХ ♂ аа G: Aa F1: Aа – 100 % |
| выяснение генотипа |
ОПЫТЫ ТОМАСА МОРГАНА
| сер./норм. чер./зач. P: ♀ ААBB Х ♂ ааbb G: АB аb F1: АаBb – 100 % серые с нормальными крыльями |
| 2 пары анализируемых признаков: А – серое тело; а – черное тело; В – нормальные крылья; b – зачаточные крылья. |
При проведении анализирующего скрещивания самки (F1) из первого поколения с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% – серые длиннокрылые и 41,5% – черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% – черные длиннокрылые и 8,5% – серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов – АВ и аb, а отцовский – один тип – аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АаВb и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb.
Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток – мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Причиной возникновения новых гамет является обмен участками гомологичных хромосом, или кроссинговер.
АНАЛИЗИРУЮЩЕЕ СКРЕЩИВАНИЕ
ОПЫТ №1. ПОЛНОЕ СЦЕПЛЕНИЕ
(из F1 берется самец – АаBb)
| Следовательно, дигетерозиготный самец(АаBb)дает два сорта гамет: ● АB; ● аb |
| P: ♀ ааbb Х ♂ (F1)АаBb G: аb? F2: АаBb – 50% – серые с нормальными крыльями; ааbb – 50% – черные с зачаточными крыльями. |
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза) (рис. 9).

Рис. 9. 1 – механизм цис-фазы (некроссоверные гаметы);
2 – механизм транс-фазы (некроссоверные гаметы).
ОПЫТ №2. НЕПОЛНОЕ СЦЕПЛЕНИЕ. КРОССИНГОВЕР.
(из F1 берется самка – АаBb (рис. 10))
| P: ♀(F1) АаBb Х ♂ ааbb G:? аb F2: АаBb – 41,5% – серые с нормальными крыльями; ааbb – 41,5% – черные с зачаточными крыльями; ааBb – 8,5% – черные с нормальными крыльями Ааbb – 8,5% – серые с зачаточными крыльями |
| 17% – КРОС-СОВЕРНЫЕ ОСОБИ |
| Следовательно, дигетерозиготная самка (АаBb) дает четыре сорта гамет: ● АB – 41,5%; ● аb – 41,5%; ● аB – 8,5% ● Аb – 8,5% |
| 17% – КРОССОВЕРНЫЕ ГАМЕТЫ |
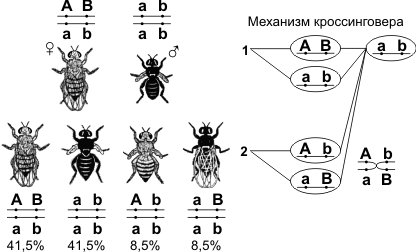
Рис. 10. 1 – некроссоверные гаметы; 2 – кроссоверные гаметы.
Таким образом, у самки, как и у самца, гены А, B находятся в одной хромосоме. Но у самца они располагаются ближе друг к другу и удерживаются сильнее (ПОЛНОЕ СЦЕПЛЕНИЕ). У самки гены А и В находятся на некотором расстоянии друг от друга, сцеплены более слабо друг с другом, чем у самца, поэтому в 17% наблюдается кроссинговер (НЕПОЛНОЕ СЦЕПЛЕНИЕ).
Расстояние между генами измеряется в морганидах – условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозигота АаВb образует четыре типа гамет (АВ, Аb, аВ и аb) в равных количествах, то такая же дигетерозигота в случае сцепления генов А и В образует только два типа гамет – АВ и аb – тоже в равных количествах. Это некроссоверные гаметы – гаметы, в процессе образования которых кроссинговер не произошел. Последние повторяют комбинацию генов в хромосоме родителя.
 образуются гаметы:
образуются гаметы: 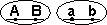
Нерекомбинанты – гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Однако полное сцепление генов наблюдается достаточно редко, и в потомстве обычно бывают представлены все четыре фенотипа. Таким образом, и в этом случае при дигибридном скрещивании образуются новые сочетания признаков – рекомбинантные фенотипы. Было установлено, что кроме обычных гамет в этом случае возникают и другие – Аb и аВ – кроссоверные гаметы, то есть появляются гаметы с новым сочетанием (комбинацией) аллелей, отличающимися от родительской гаметы.
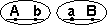 образуются гаметы:
образуются гаметы: 
Причиной возникновения новых гамет является обмен участками гомологичных хромосом, или кроссинговер. Если особи с новыми генными комбинациями (рекомбинанты – гибридные особи, имеющие иное сочетание признаков, чем у родителей) встречаются в потомстве реже, чем особи с родительскими фенотипами, то это верный признак сцепленности соответствующих генов.
После опытов Томаса Моргана мушка дрозофила стала излюбленным объектом исследования генетиков, потому что:
1) неприхотлива в содержании (легко размножается в лабораторных условиях при температуре 25°Ϲ);
2) выраженный половой диморфизм;
3) очень плодовита (большое число потомков);
4) быстрая смена поколений;
5) только 8 хромосом (небольшое число групп сцепления), но огромное количество альтернативных признаков.
Группа сцепления – гены, локализованные в одной хромосоме и наследующиеся совместно. Число групп сцепления соответствует гаплоидному набору хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены.
У человека количество групп сцепления зависит от пола – женский пол – 23 группы сцепления, у мужчин – 24 группы сцепления, поскольку две половые хромосомы (Х и Y) не являются гомологичными и несут различный набор генов.
Сцепленное с полом наследование. Примеры расписать.
Наследование сцепленное с полом.
Половые хромосомы Х и Y содержат большое количество генов. Наследование определяемых ими признаков называют наследованием, сцепленным с полом, а локализацию генов в половых хромосомах называют сцеплением генов
с полом.Например, Х-хромосома человека содержит доминантный ген Н,
пределяющий свертывание крови. У человека, рецессивно гомозиготного по этому признаку, развивается тяжелое заболевание гемофилия, при котором кровь не сворачивается и человек может погибнуть от малейшего повреждения сосудов. Так как в клетках женщин две Х-хромосомы, то наличие в одной из них гена h не влечет за собой заболевания, так как во второй из них присутствует доминантный ген Н. В клетках мужчин есть только одна Х-хромосома. Если в ней присутствует ген h, то у мужчины разовьется гемофилия, так как Y-хромосома не гомологична Х-хромосоме и в ней нет гена Н или h.
|
|
|

Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...
© cyberpedia.su 2017-2026 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

