Строение и функции ДНК у прокариот, вирусов и эукариот
Сначала мы рассмотрим строение и функции ДНК. Две полинуклеотидные нити удерживаются между собой многократно повторяющимися водородными связями, образующимися между азотистыми основаниями. В паре "аденин - тимин" присутствуют две, а в паре "гуанин - цитозин" - три водородные связи.
Консервативное соответствие пуриновых и пиримидиновых оснований было открыто Э. Чаргаффом и получило название принципа комплементарности. В отдельно взятой цепи нуклеотиды связаны между собой фосфодиэфирными связями, формирующимися между пентозой и остатком ортофосфорной кислоты рядом расположенных нуклеотидов. Спиральный вид обеих цепей поддерживается водородными связями, возникающими между атомами водорода и кислорода, находящимися в составе нуклеотидов. Высшая – третичная структура (суперспираль) - характерна для ядерной ДНК эукариотических клеток. В таком виде она присутствует в хроматине. Однако бактерии и ДНК-содержащие вирусы имеют дезоксирибонуклеиновую кислоту, не связанную с белками. Она представлена кольцеобразной формой и называется плазмидой. Такой же вид имеет ДНК митохондрий и хлоропластов – органелл растительных и животных клеток.
ДНК, имеющая конфигурацию суперспирали, находится в ядре. Функция - сохранение и передача наследственной информации.
ДНК, находящаяся в митохондрии или в хлоропласте, имеет кольцевую конфигурацию (плазмида). Функция - локальная передача наследственной информации.
Нуклеиновые кислоты вирусов могут иметь вид как одно-, так и двухнитевых спиралей или колец. Согласно классификации Д.Балтимора, эти объекты микромира содержат молекулы ДНК, состоящие из одной или двух цепей. К первой группе относятся возбудители герпеса и аденовирусы, а во вторую входят, например, парвовирусы.
Функции ДНК вирусов заключаются в проникновении собственной наследственной информации в клетку, проведении реакций репликации молекул вирусной нуклеиновой кислоты и сборке белковых частиц в рибосомах клетки-хозяина. В итоге весь клеточный метаболизм оказывается полностью подчинен паразитам, которые, стремительно размножаясь, приводят клетку к гибели.
Репликация ДНК
Репликация (возобновление) – процесс синтеза дочерней молекулы ДНК на матрице родительской молекулы ДНК. Наблюдения показали, что репликация начинается не с конца линейной молекулы, а с образования репликативного глаза, где две цепи родительской ДНК отделяются друг от друга, и в местах расхождения полинуклеотидных цепочек начинается синтез дочерних цепей. Это приводит к появлению в молекуле ДНК петли с виде участка, расплетенного на две отдельные нити.

Участок нуклеиновой кислоты, с которого начинается репликация называется точкой инициации репликации или ориджином. Этот участок содержит последовательность, состоящую из 300 нуклеотидов. Область расхождения двойной спирали ДНК на одинарных полинуклеотидные цепочки в зонах репликации называют репликативной вилкой. Постепенное разъединение цепей молекулы ДНК происходит при разрыве водородных связей между пуринами и пиримидинами. Репликация может происходить в одном или двух направлениях. При однонаправленной репликации вдоль ДНК движется одна реплекационная вилка. При двунаправленной от точки начала в противоположные стороны расходятся две вилки. На определенном расстоянии от точки инициации молекула ДНК содержит участок, в котором репликация останавливается (точки терминации репликации). Участок от точки инициации до точки ее окончания – репликон – единица репликации. Для осуществления репликации цепи материнской ДНК должны быть отделены друг от друга, чтобы стать матрицами, на которых будут синтезироваться комплементарные цепи дочерних молекул. Такое разделение комплементарных полинуклеотидных цепочек происходит с помощью фермента геликазы. Геликаза, разделяя спирально закрученные цепи материнской ДНК, вызывает появление супервитков (те сильную спирализацию молекулы ДНК) перед репликационной вилкой. Супервитки возникают потому, что при расхождении каждых 10 пар нуклеотидов, образующих один виток спирали, материнская ДНК должна совершить один полный оборот вокруг своей оси. Следовательно, для продвижения репликационной вилки вперед вся молекула перед ней должна была бы быстро вращаться. Такое вращение потребовало бы большой затраты энергии. В действительности же вращения не происходит благодаря действию особого фермента – ДНК-топоизомеразы. Топоизомераза разрывает одну из цепей ДНК и совершает оборот вокруг второй (целой) цепи как вокруг оси вращения. 
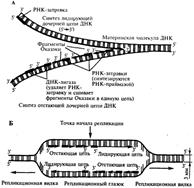
Это устраняет супервитки и ослабляет накопившиеся напряжение в двойной спирали ДНК. После снятия избыточного напряжения фермент восстанавливает разорванную цепь. К высвобождающимся водородным связям нуклеотидных последовательностей разделенных родительских цепей присоединяются свободные нуклеотиды из нуклеоплазмы, где они присутствуют в виде дезоксирибонуклеозидгрифосфатов: дАТФ, дГТФ, дЦТФ, дТТФ. Комплементарный нуклеозидтрифосфат образует водородные связи с определенным основанием материнской цепи ДНК. Затем при участии фермента ДНК-полимеразы он связывается фосфодиэфирной связью с предшествующим нуклеотидом вновь синтезируемой цепи, отдавая при этом неорганический пирофосфат. Поскольку ДНК-полимераза присоединяет очередной нуклеотид к ОН-группе в 3'-положении предшествующего нуклеотида, цепь постепенно удлиняется на ее 3'-конце. Особенностью ДНК-полимеразы является ее неспособность начать синтез новой полинуклеотидной цепи путем простого связывания двух нуклеозидтрифосфатов: необходим 3'-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинук-леотиднуй цепь называют затравкой или праймером. Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы (рис. 3.11). Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3'-ОН-конец. Способность ДНК-полимеразы осуществлять сборку полинуклеотида в направлении от 5'- к 3' -концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Действительно, если на одной из матриц (3' → 5') сборка новой цепи происходит непрерывно от 5'- к 3'-концу и она постепенно удлиняется на 3'-конце, то другая цепь, синтезируемая на матрице (5' → 3'), должна была бы расти от 3'- к 5'-концу. Это противоречит направлению действия фермента ДНК-полимеразы. В настоящее время установлено, что синтез второй цепи ДНК осуществляется короткими фрагментами (фрагменты Оказаки) также в направлении от 5'- к 3'-концу (по типу шитья «назад иголкой»). У прокариот фрагменты Оказаки содержат от 1000 до 2000 нуклеотидов, у эукариот они значительно короче (от 100 до 200 нуклеотидов). Синтезу каждого такого фрагмента предшествует образование РНК-затравки длиной около 10 нуклеотидов. Вновь образованный фрагмент с помощью фермента ДНК-лигазы соединяется с предшествующим фрагментом после удаления его РНК-затравки (рис. 3.12, А). В связи с указанными особенностями репликационная вилка является асимметричной. Из двух синтезируемых дочерних цепей одна строится непрерывно, ее синтез идет быстрее и эту цепь называют лидирующей. Синтез другой цепи идет медленнее, так как она собирается из отдельных фрагментов, требующих образования, а затем удаления РНК-затравки. Поэтому такую цепь называют запаздывающей (отстающей). Хотя отдельные фрагменты образуются в направлении 5' → 3', в целом эта цепь растет в направлении 3' → 5' (рис. 3.12, А). В виду того, что от локуса ori как правило начинаются две репликационные вилки, идущие в противоположных направлениях, синтез лидирующих цепей в них идет на разных цепях материнской ДНК (рис 3.12, Б). Конечным результатом процесса репликации является образование двух молекул ДНК, нуклеотидная последовательность которых идентична таковой в материнской двойной спирали ДНК. Рассмотренная последовательность событий, происходящих в ходе репликативного синтеза, предполагает участие целой системы ферментов: геликазы, топоизомеразы, дестабилизирующих белков, ДНК-полимеразы и других, совместно действующих в области репликационной вилки (рис 3.13). Репликация ДНК у про- и эукариот в основных чертах протекает сходно, однако, скорость синтеза у эукариот (около 100 нуклеотидов/с) на порядок ниже, чем у прокариот (1000 нуклеотидов/с). Причиной этого может быть образование ДНК эукариот достаточно прочных соединений с белками (см. гл 3.5.2.), что затрудняет ее деспирали-зацию, необходимую для осуществления репликативного синтеза. Фрагмент ДНК от точки начала репликации до точки ее окончания образует единицу репликации — репликон. Однажды начавшись в точке начала (локус on), репликация продолжается до тех пор, пока весь репликон не будет дуплицирован. Кольцевые молекулы ДНК прокариотических клеток имеют один локус on и представляют собой целиком отдельные репликоны. Эукариотические хромосомы содержат большое число репликонов. В связи с этим удвоение молекулы ДНК, расположенной вдоль эукариотической хромосомы, начинается в нескольких точках. В разных репликонах удвоение может идти в разное время или одновременно.
Детерминация. Типы детерминации. Примеры.
Детерминация представляет собой важное событие, в результате которого клетки с одинаковым набором генов начинают различаться по своим внешним признакам, или фенотипу.выдающийся швейцарский генетик Э. Хадорн использовал плодовую мушку дрозофилу. Известно, что в развитии высших насекомых, в том числе мух, происходит интересное разделение клеток по их функциям. Клетки одного типа начинают дифференцироваться с первых этапов эмбрионального развития, из них образуется тело личинки насекомого со всеми его органами. Клетки другого типа обособлены, они составляют так называемые имагинальные диски или зачатки взрослых органов. Хотя клетки имагинальных дисков находятся в контакте с соседними дифференцирующимися клетками, они находятся в эмбриональном состоянии в течение всего личиночного периода. В это время они делятся. В процессе
Метаморфоза значительная часть личиночных органов рассасывается (или лизируется). Одновременно с этим клетки имагинальных дисков утрачивают свое эмбриональное состояние, они дифференцируются, превращаясь в специализированные ткани имаго (взрослой мухи). Из каждого диска образуется отдельная часть тела насекомого. Например, для каждой из шести будущих ног существует отдельный диск, голова образуется из трех пар дисков.
Имагинальные диски можно извлечь из тела личинки и пересадить в полость тела другой личинки. Когда личинкахозяин превращается в куколку, трансплантант дифференцируется в соответствующий орган. Например, если трансплантирован глазной имагинальный диск, в брюшке личинки-хозяинаразвивается полностью сформировавшийся глаз.
В результате многих опытов Э. Хадорн обнаружил, что каждый имагинальный диск представляет собой своего рода мозаику из различных групп клеток, например, из одних участков мужского генитального диска образуется семяизвергательный канал, из других - различные элементы мужского полового члена, из третьих - анальные пластинки и задняя кишка. Следовательно, будущее разнообразие клеток детерминировано уже на личиночной стадии.
Неожиданные результаты были получены после трансплантации имагинальных дисков сразу во взрослых мух. Клетки неограниченно делились и разрастались. Если бы развитие этихимагинальных дисков происходило в нормальной личинке, то эти клетки прекратили бы деление с началом метаморфоза: под влиянием гормона насекомых экдистерона они бы начали дифференцироваться в такие структуры взрослых особей, как щетинки, волоски, коготки и т.д. Неограниченный рост этих клеток, пересаженных в брюшко взрослых мух, продолжался более 6 лет. Поскольку муха дрозофила живет около месяца, размножающиеся клетки пересаживали в новую муху через каждые 2 недели. При этом клетки перенесли свыше 160 пересадок. Хотя трансплантанты жили во взрослых мухах годами, они сохранили свой исходный эмбриональный характер и не дифференцировались. Если же извлечь немного трансплантанта и ввести в личинку, эти клетки претерпевали метаморфоз и нормально дифференцировались в структуры взрослого организма. При этом, если несколько лет назад для трансплантации были взяты крыловые имагинальные диски, при обратной трансплантации также формировалось крыло. Таким образом, состояние детерминации может воспроизводиться длительное время без каких-либоизменений. Очень продолжительное время это свойство детерминированных клеток передается благодарясвоего рода “клеточной наследственности” или определенного состоянияядерно-цитоплазматическихотношений (или наличия в цитоплазме эпигенетических факторов).
В некоторых случаях нормальное состояние детерминированности в опытах Э. Хадорна резко изменялось.
Очень редко, из клеток генитального имагинального диска после длительного размножения в брюшке имаго, формировались органы головы или конечности, т.е. клетки больше не дифференцировались в соответствие с детерминацией их предков - произошла трансдетерминация. Вновь приобретенное трансдетерминированное состояние в дальнейшем также передается за счет клеточной наследственности очень продолжительное время.
Явление трансдетерминации, также как и результаты трансплантации ядер, полученные Дж. Гердоном свидетельствуют, что в основе дифференцировки не лежит необратимое состояние генов, тем более их потеря.
Очевидно также и то, что в основе любого детерминированного состояния лежит сбалансированная система ядерноцитоплазматических отношений. Как может сформироваться такая система лучше всего показывают результаты самого раннего развития.



Предмет, задачи и методы генетики.
Предмет: впервые термин генетика был введен У. Бэтсооном в 1906г. Происходит от слова Происходит от слова genesis, что означает «происхождение». Генетика изучает два неразрывных свойства живых организмов: наследственность и изменчивость, а также методы управления ими. Именно наследственность и изменчивость являются предметом генетики. Изменчивость – это разнообразие. Свойства наследственности и изменчивости прослеживаются в пределах отдельных видов. Пример – человек. Разнообразие людей. Варьирует морфология. В то же время каждый человек знает те черты, которыми напоминает своих родителей. Именно благодаря механизму наследования каждый индивидуум имеет черты сходства с предками. Именно поэтому, каждый человек появляется в результате слияния гамет и перекомбинации генов в длительном ряду поколений, дети никогда не повторяют своих родителей. Вообще невозможно найти двух идентичных людей. Чрезвычайно похожи только однояйцовые близнецы, и то лишь потому, что появились в результате вегетативного размножения – деления одной и той же оплодотворенной клетки. Г.Мендель в 1865г сформировал законы наследования дискретных факторов, или генов, как их теперь называют.
Задачи генетики вытекают из установленных общих закономерностей наследственности и изменчивости. К этим задачам относят исследования: 1) механизмов хранения и передачи информации от родительских форм дочерним. 2) механизм реализации этой информации в виде признаков и свойств организмов в процессе их индивидуального развития под контролем генов и влиянием условий внешней среды. 3) типов, причин и механизмов изменчивости всех живых существ. 4) взаимосвязи процессов наследственности, изменчивости и отбора как движущих факторов эволюции органического мира.
Генетика является так же основой для решения ряда важнейших практических задач. К ним относятся: 1)Выбор наиболее эффективных типов гибридизации и способов отбора. 2) управление развитием наследственных признаков с целью получения наиболее значимых для человека результатов. 3) искусственное получение наследственно измененных форм живых организмов. 4) разработка мероприятий по защите живой природы от вредных мутагенов внешней среды и методов борьбы с наследственными болезнями человека, вредителями сельскохозяйственных растений и животных. 5) разработка методов генетической инженерии с целью получения высокоэффективных продуцентов биологически активных соединений, а также для создания принципиально новых технологий в селекции микроорганизмов, растений животных.
Методы: 1) Гибридологический метод – представляет собой специфический метод генетики. Он в значительной степени совпадает с методом генетического анализа. Метод гибрид.анализа заключается в гибридизации и последующем учете расщепления, в законченной форме был предложен Г.Менделем. Им были сформулированы непреложные правила, которым следуют все генетики: 1.скрещиваемые организмы должны принадлежать одному виду. 2. Скрещиваемые организмы должны четко различаться по отдельным признакам. 3. Признаки должны быть константны – воспроизводиться из поколения в поколение при скрещивании в пределах линии (родительской формы). 4. Необходимы характеристика и количественный учет всех классов расщепления, если оно наблюдается у гибридов первого поколения и последующих поколений.
2) Математический метод – Мендель применил количественный подход к изучению результатов скрещивания, а также, что не менее важно, к построению гипотез, объясняющих полученный результаты. С тех пор сравнение количественных данных эксперимента с теоретически ожидаемыми стало неотъемлемой частью генетического анализа.
3) Цитологический метод – используется для изучения клетки как основной единицы живой материи. Исследование строения хромосом вместе с гибридологическим анализом лежит в основе цитогенетики.
Генетика активно использует и методы других смежных наук. Методы химии и биохимии, физики (оптические, седиментационные, методы меченых атомов для маркирования и идентификации различных классов макромолекул). Наиболее широко физические, химические и физико-химические методы применяются в молекулярной генетике и генной инженерии. Генетики, работающие с различными объектами, не могут обойтись и без методов медицины, зоологии, ботаники, микробиологии и других дисциплин. В то же время все большая связь с эволюционной теорией повышает значение для генетики сравнительного анализа.





